Physicochemical Properties
| Molecular Formula | C46H77NO17 |
| Molecular Weight | 916.1001 |
| Exact Mass | 915.519 |
| Elemental Analysis | C, 60.31; H, 8.47; N, 1.53; O, 29.69 |
| CAS # | 1401-69-0 |
| Related CAS # | Tylosin tartrate;74610-55-2;Tylosin phosphate;1405-53-4;Tylosin-d3 |
| PubChem CID | 5280440 |
| Appearance | White to light yellow solid powder |
| Density | 1.2±0.1 g/cm3 |
| Boiling Point | 980.7±65.0 °C at 760 mmHg |
| Melting Point | 18-132ºC |
| Flash Point | 546.9±34.3 °C |
| Vapour Pressure | 0.0±0.6 mmHg at 25°C |
| Index of Refraction | 1.549 |
| LogP | 3.27 |
| Hydrogen Bond Donor Count | 5 |
| Hydrogen Bond Acceptor Count | 18 |
| Rotatable Bond Count | 13 |
| Heavy Atom Count | 64 |
| Complexity | 1560 |
| Defined Atom Stereocenter Count | 21 |
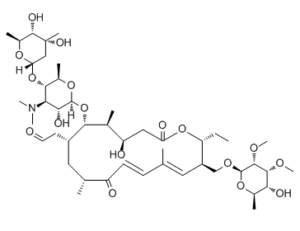
| SMILES | O1[C@]([H])(C([H])([H])[H])C([H])([C@@]([H])(C([H])(C1([H])OC1([H])C([H])(C([H])([H])[H])[C@@]([H])(C([H])([H])C(=O)OC([H])(C([H])([H])C([H])([H])[H])C([H])(C([H])=C(C([H])([H])[H])C([H])=C([H])C(C([H])(C([H])([H])[H])C([H])([H])[C@]1([H])C([H])([H])C([H])=O)=O)C([H])([H])OC1([H])C([H])([C@@]([H])(C([H])(C([H])(C([H])([H])[H])O1)O[H])OC([H])([H])[H])OC([H])([H])[H])O[H])O[H])N(C([H])([H])[H])C([H])([H])[H])OC1([H])C([H])([H])C(C([H])([H])[H])([C@]([H])([C@]([H])(C([H])([H])[H])O1)O[H])O[H] |c:45,52| |
| InChi Key | WBPYTXDJUQJLPQ-VMXQISHHSA-N |
| InChi Code | InChI=1S/C46H77NO17/c1-13-33-30(22-58-45-42(57-12)41(56-11)37(52)26(5)60-45)18-23(2)14-15-31(49)24(3)19-29(16-17-48)39(25(4)32(50)20-34(51)62-33)64-44-38(53)36(47(9)10)40(27(6)61-44)63-35-21-46(8,55)43(54)28(7)59-35/h14-15,17-18,24-30,32-33,35-45,50,52-55H,13,16,19-22H2,1-12H3/b15-14+,23-18+/t24-,25+,26-,27-,28+,29+,30-,32-,33-,35+,36-,37-,38-,39-,40-,41-,42-,43+,44+,45-,46-/m1/s1 |
| Chemical Name | 2-((4R,5S,6S,7R,9R,11E,13E,15R,16R)-6-(((2R,3R,4R,5S,6R)-5-(((2S,4R,5S,6S)-4,5-dihydroxy-4,6-dimethyltetrahydro-2H-pyran-2-yl)oxy)-4-(dimethylamino)-3-hydroxy-6-methyltetrahydro-2H-pyran-2-yl)oxy)-16-ethyl-4-hydroxy-15-((((2R,3R,4R,5R,6R)-5-hydroxy-3,4-dimethoxy-6-methyltetrahydro-2H-pyran-2-yl)oxy)methyl)-5,9,13-trimethyl-2,10-dioxooxacyclohexadeca-11,13-dien-7-yl)acetaldehyde |
| Synonyms | Fradizine; Tylocine; Tylosin; Tylosin A; Tilosina; Tylan; Vubityl 200. |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets | Macrolide |
| ln Vitro |
Tylosin binds to the bacterial ribosomal 50S subunit's 23S rRNA to produce antibacterial effects [1]. With minimum inhibitory concentrations (MICs) of 64 μg/mL, 32 μg/mL, 512 μg/mL, and 1 μg/mL for M. haemolytica 11935, P. multocida 4407, E. coli ATCC 25922 and E. coli AS19rlmAI, respectively, tylosin also inhibits the growth of Gram-negative strains[3]. |
| ln Vivo | Animals treated with lipopolysaccharide (LPS) showed an increase in IL-10 and a general suppression of elevated TNF-α and IL-1β levels when given tylosin (10–500 mg/kg; s.c.)[4]. |
| Animal Protocol |
Animal Model: Balb/C mice (2-3 months old, 20-25 g)[4] Dosage: 10 mg/kg, 100 mg/kg, 500 mg/kg Administration: Subcutaneous injection Result: raised the levels of IL-10 in mice treated with 250 µg of LPS, but decreased the elevated levels of TNF-α and IL-1β. |
| ADME/Pharmacokinetics |
Absorption, Distribution and Excretion The pharmacokinetics and oral bioavailability of tylosin tartrate and tylosin phosphate were carried out in broiler chickens according to a principle of single dose, random, parallel design. The two formulations of tylosin were given orally and intravenously at a dose level of 10 mg/kg b.w to chicken after an overnight fasting (n = 10 chickens/group). Serial blood samples were collected at different time points up to 24 hr postdrug administration. A high performance liquid chromatography method was used for the determination of tylosin concentrations in chicken plasma. The tylosin plasma concentration's time plot of each chicken was analyzed by the 3P97 software. The pharmacokinetics of tylosin was best described by a one-compartmental open model 1st absorption after oral administration. After intravenous administration the pharmacokinetics of tylosin was best described by a two-compartmental open model, and there were no significant differences between tylosin tartrate and tylosin phosphate. After oral administration, there were significant differences in the Cmax (0.18 + or - 0.01, 0.44 + or - 0.09) and AUC (0.82 + or - 0.05, 1.57 + or - 0.25) between tylosin phosphate and tylosin tartrate. The calculated oral bioavailability (F) of tylosin tartrate and tylosin phosphate were 25.78% and 13.73%, respectively. Above all, we can reasonably conclude that, the absorption of tylosin tartrate is better than tylosin phosphate after oral administration. /MILK/ The aim of this study is to determine the pharmacokinetics of tylosin and tilmicosin in serum and milk in healthy Holstein breed cows (n = 12) and reevaluate the amount of residue in milk. Following the intramuscular administration of tylosin, the maximum concentrations (C max) in serum and milk were found to be 1.30 + or - 0.24 and 4.55 + or - 0.23 ug/mL, the time required to reach the peak concentration (t max) was found to be 2nd and 4th hour, and elimination half-live were found to be 20.46 + or - 2.08 and 26.36 + or - 5.55 hour, respectively. Following the subcutaneous administration of tilmicosin, the C max in serum and milk were found to be 0.86 + or - 0.20 and 20.16 + or - 1.13 ug/mL, the t max was found to be 1st and 8th hr, and the elimination half life were found to be 29.94 + or - 6.65 and 43.02 + or - 5.18 hr, respectively. AUCmilk/AUCserum and C max-milk/C max-serum rates, which are indicators for determining the rate of drugs that pass into milk, were, respectively, calculated as 5.01 + or - 0.72 and 3.61 + or - 0.69 for tylosin and 23.91 + or - 6.38 and 20.16 + or - 1.13 for tilmicosin. In conclusion, it may be stated that milk concentration of tylosin after parenteral administration is higher than expected like tilmicosin and needs more withdrawal period for milk than reported. Biological availability and pharmacokinetic properties of tylosin were determined in broiler chickens after oral and iv administration at a dose of 10 mg/kg. The calculated bioavailability--F%, by comparing AUC values--oral and AUC--iv, ranged from 30%-34%. After intravenous injection tylosin was rapidly distributed in the organism, showing elimination half-life values of 0.52 hr and distribution volume (Vd) of 0.69 L/kg, at a clearance rate (Cl) of 5.30 +/- 0.59 mL/min/kg. After oral administration, tylosin has a similar distribution volume (Vd = 0.85 L/kg), while the elimination half-life of 2.07 hr was four times bigger than after iv administration at Cl = 4.40 +/- 0.27 mL/min/kg. The obtained value tmax = 1.5 hr for tylosin after oral administration indicates that using this antibiotic with drinking water in broiler chickens is the method of choice. However, a relatively low value Cmax = 1.2 micrograms/ml after oral administration of tylosin shows that dosing of this antibiotic in broiler chickens should be higher than in other food producing animals. /MILK/ Antibiotic residues in milk above tolerance levels interfere with dairy product processing and pose potential health risks to consumers. Residue avoidance programmes include, among other components, the observance of withdrawal times indicated in label instructions. Persistence of antibiotics in milk following treatment is influenced by drug, dosage, route of administration, body weight and mammary gland health status. Compositional changes that take place during intramammary infection (IMI) can affect antibiotic excretion in milk, thus modifying milk withdrawal time. The objectives of this study were to validate sensitivity and specificity of a qualitative microbiological method (Charm AIM-96) to detect tylosin in bovine composite milk and to determine the influence of subclinical IMI in tylosin excretion following intramuscular administration. For test validation, two groups of approximately 120 cows were used; one received a single intramuscular injection of tylosin tartrate at a dose of 20 mg/kg, while the other group remained as untreated control. Test sensitivity and specificity were 100% and 94.1% respectively. To determine the influence of subclinical IMI in tylosin excretion, two groups of seven cows, one with somatic cell counts (SCC) < or =250 000 cells/ml and the other with SCC > or =900 000, were administered a single intramuscular injection of tylosin tartrate at a dose of 20 mg/kg. Milk samples were obtained every 12 h for 10 days following treatment. Milk tylosin excretion averaged between 5 and 9 days for cows with low and high SCC respectively (P < 0.0001). Compositional changes in cows with high SCC most likely affect the pharmacokinetic characteristics of tylosin, extending the presence of the antibiotic in milk, thus influencing milk withdrawal times. For more Absorption, Distribution and Excretion (Complete) data for TYLOSIN (10 total), please visit the HSDB record page. Metabolism / Metabolites The tylosin-biosynthetic (tyl) gene cluster of Streptomyces fradiae contains ancillary genes that encode functions normally associated with primary metabolism. These can be disrupted without loss of viability, since equivalent genes (presumably used for 'housekeeping' purposes) are also present elsewhere in the genome. The tyl cluster also contains two genes that encode products unlike any proteins in the databases. Two ancillary genes, metF (encoding N5,N10-methylenetetrahydrofolate reductase) and metK, encoding S-adenosylmethionine synthase, flank one of the 'unknown' genes (orf9) in the tyl cluster. In a strain of S. fradiae in which all three of these genes were disrupted, tylosin production was reduced, although this effect was obscured in media supplemented with glycine betaine which can donate methyl groups to the tetrahydrofolate pool. Apparently, one consequence of the recruitment of ancillary genes into the tyl cluster is enhanced capacity for transmethylation during secondary metabolism. Studies on the susceptibility of pathogenic Nocardia to macrolide antibiotics, chalcomycin and tylosin, showed that most of the Nocardia species examined were highly resistant to both antibiotics, although N. nova was moderately susceptible. N. asteroides IFM 0339 converted these macrolides into inactive metabolites by glycosylation at 2'-OH or glycosylation and reduction of the 20-formyl group. The structures of the metabolites were determined from NMR and MS data to be 2'-[O-(beta-D-glucopyranosyl)]chalcomycin (2), 2'-[O-(beta-D-glucopyranosyl)]tylosin (5) and 20-dihydro-2'-[O-(beta-D-glucopyranosyl)]tylosin (4). Tylosin is produced by Streptomyces fradiae via a combination of polyketide metabolism and synthesis of three deoxyhexose sugars, of which mycaminose is the first to be added to the polyketide aglycone, tylactone (protylonolide). Previously, disruption of the gene (tylMII) encoding attachment of mycaminose to the aglycone unexpectedly abolished accumulation of the latter, raising the possibility of a link between polyketide metabolism and deoxyhexose biosynthesis in S. fradiae. However, at that time, it was not possible to eliminate an alternative explanation, namely, that downstream effects on the expression of other genes, not involved in mycaminose metabolism, might have contributed to this phenomenon. Here, it is shown that disruption of any of the four genes (tylMI--III and tylB) specifically involved in mycaminose biosynthesis elicits a similar response, confirming that production of mycaminosyl-tylactone directly influences polyketide metabolism in S. fradiae. Under similar conditions, when mycaminose biosynthesis was specifically blocked by gene disruption, accumulation of tylactone could be restored by exogenous addition of glycosylated tylosin precursors. Moreover, certain other macrolides, not of the tylosin pathway, were also found to elicit qualitatively similar effects. Comparison of the structures of stimulatory macrolides will facilitate studies of the stimulatory mechanism. Three glycosyltransferases are involved in tylosin biosynthesis in Streptomyces fradiae. The first sugar to be added to the polyketide aglycone (tylactone) is mycaminose and the gene encoding mycaminosyltransferase is orf2* (tylM2). However, targeted disruption of orf2* did not lead to the accumulation of tylactone under conditions that normally favor tylosin production; instead, the synthesis of tylactone was virtually abolished. This may, in part, have resulted from a polar effect on the expression of genes downstream of orf2*, particularly orf4* (ccr) which encodes crotonyl-CoA reductase, an enzyme that supplies 4-carbon extender units for polyketide metabolism. However, that cannot be the entire explanation, since tylosin production was restored at about 10% of the wild-type level when orf2* was re-introduced into the disrupted strain. When glycosylated precursors of tylosin were fed to the disrupted strain, they were converted to tylosin, confirming that two of the three glycosyltransferase activities associated with tylosin biosynthesis were still intact. Interestingly, however, tylactone also accumulated under such conditions and, to a much lesser extent, when tylosin was added to similar fermentations. It is concluded that glycosylated macrolides exert a pronounced positive effect on polyketide metabolism in S. fradiae. For more Metabolism/Metabolites (Complete) data for TYLOSIN (6 total), please visit the HSDB record page. Biological Half-Life Biological availability and pharmacokinetic properties of tylosin were determined in broiler chickens after oral and iv administration at a dose of 10 mg/kg. ... After intravenous injection, tylosin ... /had an/ elimination half-life value of 0.52 .. . After oral administration, tylosin /had an / elimination half-life of 2.07 hr ... . The elimination half-life of tylosin is reportedly 54 minutes in small animals, 139 minutes in newborn calves, and 64 minutes in calves 2 months of age or older. |
| References |
[1]. Resistance to the macrolide antibiotic tylosin is conferred by single methylations at 23S rRNA nucleotides G748 and A2058 acting in synergy. Proc Natl Acad Sci U S A. 2002 Nov 12; 99(23): 14658-14663. [2]. In Vitro Evaluation of the Effects of Tylosin on the Composition and Metabolism of Canine Fecal Microbiota. Animals (Basel). 2020 Jan; 10(1): 98. [3]. Inhibition of Protein Synthesis on the Ribosome by Tildipirosin Compared with Other Veterinary Macrolides. Antimicrob Agents Chemother. 2012 Nov; 56(11): 6033-6036. [4]. Effects of tylosin on serum cytokine levels in healthy and lipopolysaccharide-treated mice. Acta Vet Hung. 2010 Mar;58(1):75-81. |
| Additional Infomation |
Tylosin is a macrolide antibiotic that is tylonolide having mono- and diglycosyl moieties attached to two of its hydroxy groups. It is found naturally as a fermentation product of Streptomyces fradiae. It has a role as a bacterial metabolite, an allergen, a xenobiotic and an environmental contaminant. It is an aldehyde, a disaccharide derivative, an enone, a leucomycin, a monosaccharide derivative and a macrolide antibiotic. It is functionally related to a tylactone. It is a conjugate base of a tylosin(1+). Tylosin is a bacteriostatic macrolide antibiotic and feed additive used in veterinary medicine. It has a broad spectrum of activity against Gram-positive organisms and a limited range of Gram-negative organisms. Tylosin is produced as a fermentation product of Streptomyces fradiae. Tylosin has been reported in Bos taurus, Streptomyces fradiae, and Streptomyces venezuelae with data available. Macrolide antibiotic obtained from cultures of Streptomyces fradiae. The drug is effective against many microorganisms in animals but not in humans. See also: Tylosin Tartrate (has salt form); Tylosin Phosphate (has salt form); Monensin; Tylosin (component of) ... View More ... Mechanism of Action The inhibition of peptide bond formation by tylosin, a 16-membered ring macrolide, was studied in a model system derived from Escherichia coli. In this cell-free system, a peptide bond is formed between puromycin (acceptor substrate) and AcPhe-tRNA (donor substrate) bound at the P-site of poly(U)-programmed ribosomes. It is shown that tylosin inhibits puromycin reaction as a slow-binding, slowly reversible inhibitor. Detailed kinetic analysis reveals that tylosin (I) reacts rapidly with complex C, i.e., the AcPhe-tRNA. poly(U).70S ribosome complex, to form the encounter complex CI, which then undergoes a slow isomerization and is converted to a tight complex, CI, inactive toward puromycin. These events are described by the scheme C + I <==> (K(i)) CI <==> (k(4), k(5)) CI. The K(i), k(4), and k(5) values are equal to 3 microM, 1.5 min(-1), and 2.5 x 10(-3) min(-1), respectively. The extremely low value of k(5) implies that the inactivation of complex C by tylosin is almost irreversible. The irreversibility of the tylosin effect on peptide bond formation is significant for the interpretation of this antibiotic's therapeutic properties; it also renders the tylosin reaction a useful tool in the study of other macrolides failing to inhibit the puromycin reaction but competing with tylosin for common binding sites on the ribosome. Thus, the tylosin reaction, in conjunction with the puromycin reaction, was applied to investigate the erythromycin mode of action. It is shown that erythromycin (Er), like tylosin, interacts with complex C according to the kinetic scheme C + Er <==> (K(er)) CEr <==> (k(6), k(7)) C*Er and forms a tight complex, CEr, which remains active toward puromycin. The determination of K(er), k(6), and k(7) enables us to classify erythromycin as a slow-binding ligand of ribosomes |
Solubility Data
| Solubility (In Vitro) |
DMSO : ≥ 100 mg/mL (~109.16 mM) H2O : ~0.67 mg/mL (~0.73 mM) |
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 2.5 mg/mL (2.73 mM) (saturation unknown) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. Solubility in Formulation 2: ≥ 2.5 mg/mL (2.73 mM) (saturation unknown) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 900 μL of 20% SBE-β-CD physiological saline solution and mix evenly. Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. Solubility in Formulation 3: ≥ 2.5 mg/mL (2.73 mM) (saturation unknown) in 10% DMSO + 90% Corn Oil (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 900 μL of corn oil and mix evenly. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.0916 mL | 5.4579 mL | 10.9158 mL | |
| 5 mM | 0.2183 mL | 1.0916 mL | 2.1832 mL | |
| 10 mM | 0.1092 mL | 0.5458 mL | 1.0916 mL |