T16Ainh-A01 is an aminophenylthiazole analog acting as a novel and potent transmembrane protein 16A (TMEM16A) inhibitor which inhibits TMEM16A-mediated chloride currents with an IC50 value of ~1 µM. TMEM16A (ANO1) functions as a calcium-activated chloride channel (CaCC). TMEM16A inhibitors are potential development candidates for drug therapy of hypertension, pain, diarrhea, and excessive mucus production.
Physicochemical Properties
| Molecular Formula | C19H20N4O3S2 |
| Molecular Weight | 416.5171 |
| Exact Mass | 416.098 |
| Elemental Analysis | C, 54.79; H, 4.84; N, 13.45; O, 11.52; S, 15.39 |
| CAS # | 552309-42-9 |
| PubChem CID | 135460621 |
| Appearance | White to off-white solid powder |
| LogP | 4.565 |
| Hydrogen Bond Donor Count | 2 |
| Hydrogen Bond Acceptor Count | 7 |
| Rotatable Bond Count | 7 |
| Heavy Atom Count | 28 |
| Complexity | 660 |
| Defined Atom Stereocenter Count | 0 |
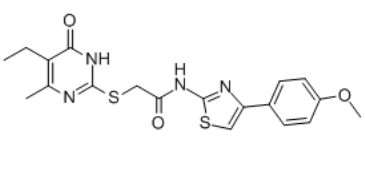
| SMILES | CCC1=C(N=C(SCC(NC2=NC(C3=CC=C(OC)C=C3)=CS2)=O)N=C1O)C |
| InChi Key | QSIYTNYMBWYHAA-UHFFFAOYSA-N |
| InChi Code | InChI=1S/C19H20N4O3S2/c1-4-14-11(2)20-18(23-17(14)25)28-10-16(24)22-19-21-15(9-27-19)12-5-7-13(26-3)8-6-12/h5-9H,4,10H2,1-3H3,(H,20,23,25)(H,21,22,24) |
| Chemical Name | 2-[(5-ethyl-4-methyl-6-oxo-1H-pyrimidin-2-yl)sulfanyl]-N-[4-(4-methoxyphenyl)-1,3-thiazol-2-yl]acetamide |
| Synonyms | T16Ainh-A01; T16Ainh A01; T16AInh-A01; 2-((5-Ethyl-4-methyl-6-oxo-1,6-dihydropyrimidin-2-yl)thio)-N-(4-(4-methoxyphenyl)thiazol-2-yl)acetamide; 2-[(5-Ethyl-1,6-dihydro-4-methyl-6-oxo-2-pyrimidinyl)thio]-N-[4-(4-methoxyphenyl)-2-thiazolyl]acetamide; T16Ainh - A01; T16Ainh-A01;; t16a(inh)-a01; 2-[(5-ethyl-6-methyl-4-oxo-1H-pyrimidin-2-yl)sulfanyl]-N-[4-(4-methoxyphenyl)-1,3-thiazol-2-yl]acetamide; T16Ainh-A-01 |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets |
TMEM16A/transmembrane protein 16A T16Ainh-A01 is an inhibitor of the calcium-activated chloride channel TMEM16A (ANO1) (IC50 ~1.1 µM). It also inhibits the related isoform TMEM16B. [2] |
| ln Vitro |
Without significantly affecting L-type Ca2+ current, T16Ainh-A01 (0.3-30 μM) greatly decreases the inward and outward components of IClCa and inhibits IClCa in RUICC [1]. At all voltages, TMEM16A chloride currents (caused by 275 nM free calcium in the pipette) are nearly entirely inhibited by T16Ainh-A01 (10 μM), suggesting a voltage-independent block mechanism [2]. T16Ainh-A01 (10 µM) fully inhibited ATP-activated TMEM16A chloride current (short-circuit current) in TMEM16A-transfected FRT cells under a transepithelial chloride gradient. [2] T16Ainh-A01 (10 µM) had little effect (<10% inhibition) on CFTR chloride conductance activated by forskolin in primary cultured human bronchial epithelial cells. [2] T16Ainh-A01 (10 µM) did not alter cytoplasmic calcium elevation stimulated by ATP or ionomycin in FRT cells. [2] In whole-cell patch clamp recordings, T16Ainh-A01 (10 µM) nearly completely inhibited TMEM16A chloride current (induced by 275 nM free Ca²⁺ in the pipette) at all voltages, indicating a voltage-independent block mechanism. [2] In human salivary gland A253 cells, T16Ainh-A01 (10 µM) strongly inhibited the characteristic outwardly rectifying CaCC current (induced by 275 nM free Ca²⁺). [2] In human intestinal T84 cells, T16Ainh-A01 (10 µM) had little effect on the total sustained CaCC current induced by ATP, but selectively inhibited (~50%) the initial, transient peak current following agonist stimulation. Similar partial inhibition of the initial peak current was observed in HT-29 intestinal cells and in primary cultures of CF human bronchial epithelial cells stimulated with UTP. [2] The inhibitory effect of T16Ainh-A01 on CaCC current in airway epithelial cells diminished when the compound was added at later time points after UTP stimulation, supporting its primary effect on the transient current component. [2] In human bronchial epithelial cells treated with IL-4 (10 ng/ml for 24 h) to upregulate TMEM16A expression, the UTP-induced initial peak current and the T16Ainh-A01-sensitive current component were markedly increased. [2] siRNA-mediated knockdown of TMEM16A in T84 cells qualitatively phenocopied the effect of T16Ainh-A01, reducing the early transient current but not the sustained CaCC current. [2] |
| ln Vivo | Interstitial cells of Cajal (ICC) isolated from the rabbit urethra exhibit Ca2+-activated Cl- currents (I ClCa) that are important for the development of urethral tone. Here, we examined if TMEM16A (ANO1) contributed to this activity by examining the effect of "new-generation" TMEM16A inhibitors, CACCinh-A01 and T16Ainh-A01, on I ClCa recorded from freshly isolated rabbit urethral ICC (RUICC) and on contractions of intact strips of rabbit urethra smooth muscle. Real-time quantitative PCR experiments demonstrated that TMEM16A was highly expressed in rabbit urethra smooth muscle, in comparison to TMEM16B and TMEM16F. Single-cell RT-PCR experiments revealed that only TMEM16A was expressed in freshly isolated RUICC. Depolarization-evoked I ClCa in isolated RUICC, recorded using voltage clamp, were inhibited by CACCinh-A01 and T16Ainh-A01 with IC50 values of 1.2 and 3.4 μM, respectively. Similarly, spontaneous transient inward currents (STICs) recorded from RUICC voltage clamped at -60 mV and spontaneous transient depolarizations (STDs), recorded in current clamp, were also inhibited by CACCinh-A01 and T16Ainh-A01. In contrast, spontaneous Ca2+ waves in isolated RUICC were only partially reduced by CACCinh-A01 and T16Ainh-A01. Finally, neurogenic contractions of strips of rabbit urethra smooth muscle (RUSM), evoked by electric field stimulation (EFS), were also significantly reduced by CACCinh-A01 and T16Ainh-A01. These data are consistent with the idea that TMEM16A is involved with CACCs in RUICC and in contraction of rabbit urethral smooth muscle[1]. |
| Enzyme Assay |
Screening Procedures[2] High throughput screening was done using an automated screening platform (Beckman) equipped with FluoStar fluorescence plate readers as described. Each well of a 96-well plate was washed three times with PBS (200 μl/wash), leaving 50 μl of PBS. Test compounds (0.5 μl) were added to each well at 25 μm final concentration. After 10 min, 96-well plates were transferred to a plate reader for fluorescence assay. Each well was assayed individually for TMEM16A-mediated I− influx by recording fluorescence continuously (400 ms/point) for 2 s (base line), then 50 μl of a 140 mm I− solution containing 200 μm ATP was added. The initial rate of I− influx was computed from fluorescence data by nonlinear regression.[2] Short Circuit Current[2] Snapwell inserts containing TMEM16A-expressing FRT cells, T84 cells, or human bronchial epithelial cells were mounted in Ussing chambers. Amiloride, CFTRinh-172, UTP, ATP, and TMEM16A inhibitors were added to the apical solution, and an equal volume of vehicle was added at the same time to the basolateral solution. Symmetrical HCO3−-buffered solutions were used for T84 cells and human bronchial epithelial cells. For FRT cells, the hemichambers were filled with the half-Cl− solution (apical) and the HCO3−-buffered solution (basolateral). Cells were bathed for a 10-min stabilization period and aerated with 95% O2/5% CO2 at 37 °C or room temperature. In some experiments, for measurement of apical chloride conductance, the basolateral membrane was permeabilized with nystatin (360 μg/ml), and a chloride gradient was applied in which the basolateral membrane was bathed with the HCO3−-buffered solution, and in the apical solution 120 mm NaCl was replaced by sodium gluconate. Short circuit current was measured using an EVC4000 Multi-Channel V/I Clamp (World Precision Instruments, Sarasota, FL) and recorded using PowerLab/8sp.[2] Patch Clamp[2] Whole cell recordings were made at room temperature on TMEM16A-expressing FRT cells and human submandibular A253 cells. The pipette solution contained 130 mm CsCl, 0.5 mm EGTA, 1 mm MgCl2, 1 mm Tris-ATP, and 10 mm HEPES (pH 7.2). The bath solution contained 140 mm N-methyl-d-glucamine-Cl, 1 mm CaCl2, 1 mm MgCl2, 10 mm glucose, and 10 mm HEPES (pH 7.4). Pipettes were pulled from borosilicate glass and had resistances of 3–5 megohms after fire polishing. Seal resistances were between 3 and 10 gigohms. After establishing the whole cell configuration, TMEM16A was activated by 100 μm ATP or by 275 nm free calcium in the pipette solution (1 mm CaCl2 added to pipette solution). Whole cell currents were elicited by applying hyperpolarizing and depolarizing voltage pulses from a holding potential of 0 mV to potentials between −100 mV and +100 mV in steps of 20 mV. Recordings were made at room temperature using an Axopatch-200B. Currents were digitized with a Digidata 1440A converter, filtered at 5 kHz, and sampled at 1 kHz.[2] Cytoplasmic Calcium Measurements[2] FRT cells in 96-well black-walled microplates were loaded with Fluo-4 NW at 48 h after plating per the manufacturer's protocol. Fluo-4 fluorescence was measured with a FLUOstar Optima fluorescence plate reader equipped with syringe pumps and custom excitation/emission filters (485/538 nm). |
| Cell Assay |
Whole-cell patch clamp recordings [1] Currents were recorded with the perforated patch configuration of the whole-cell patch clamp technique. This circumvented the problem of current rundown encountered using the conventional whole-cell configuration. The cell membrane was perforated using the antibiotic amphotericin B (600 μg/ml). Patch pipettes were initially front filled by dipping into pipette solution and then back filled with the amphotericin B containing solution. Pipettes were pulled from borosilicate glass capillary tubing (1.5 mm outer diameter, 1.17 mm inner diameter) to a tip of diameter approximately 1–1.5 μm and resistance of 2–4 MΩ. Voltage clamp commands were delivered via an Axopatch 1D patch clamp amplifier connected to a Digidata 1440A AD/DA converter interfaced to a computer running pClamp software. During experiments, the cell under study was continuously superfused with Hanks’ solution by means of a close delivery system consisting of a pipette (tip diameter 200 μm) placed approximately 300 μm away. This could be switched, with a dead-space time of around 5 s, to a solution containing a drug. All experiments were carried out at 35–37 °C.[1] Calcium imaging[1] Cells were placed in Hanks’ solution containing 100 μM Ca2+ and allowed to settle in glass bottomed Petri dishes until they had stuck down. They were then incubated in 0.4 μM fluo-4/AM for 6–8 min in the dark at room temperature before being studied at 37 °C. Cells were imaged using an iXon 887 EMCCD camera (512 × 512 pixels, pixel size 16 × 16 μm) coupled to a Nipkow spinning disk confocal head as described previously. A krypton-argon laser at 488 nm was used to excite the fluo-4, and the emitted light was detected at wavelengths >510 nm. Experiments were performed using a ×60 objective (Olympus) resulting in images of pixel size 0.266 × 0.266 μm. Images were acquired at 15 frames per second. Background fluorescence from the camera, obtained using a null frame, was subtracted from each frame to obtain “F.” F 0 was determined as the minimum fluorescence measured between oscillations under control conditions. To obtain post hoc line scan images for display in figures, a one pixel thick line was drawn centrally through the entire length of the cell and the “reslice” command in ImageJ was invoked. An example of this is illustrated in Fig. 8a which shows selected frames taken a movie stack of an RUCC during a spontaneous Ca2+ wave. The white line shown in the first frame indicates the region where the pseudo-line scan shown in Fig. 8b was obtained. Therefore, it is important to recognize that the pseudo-line scan images only represent events that occurred along the length of the line and would exclude events that occurred outside this region. ∆F/F 0 refers to the measurement of the change in Ca2+ levels from basal to peak. High-Throughput Screening (HTS) Assay: Screening for TMEM16A inhibitors was performed using FRT cells stably transfected with human TMEM16A and the halide-sensitive fluorescent protein YFP-H148Q/I152L/F46L. Cells in 96-well plates were incubated with test compounds (25 µM final concentration) for 10 minutes in a physiological chloride-containing solution. TMEM16A-mediated iodide influx was then triggered by adding an extracellular solution containing 140 mM I⁻ and 200 µM ATP. Inhibition was quantified by measuring the initial rate of decrease in YFP fluorescence caused by iodide influx. [2] Short-Circuit Current (Isc) Measurement in Epithelial Monolayers: Snapwell inserts containing cell monolayers (TMEM16A-expressing FRT, T84, or human bronchial epithelial cells) were mounted in Ussing chambers. Cells were bathed in symmetrical HCO₃⁻-buffered solutions (or a half-Cl⁻ solution for FRT cells) and aerated with 95% O₂/5% CO₂. For direct measurement of apical membrane TMEM16A conductance in FRT cells, the basolateral membrane was permeabilized with nystatin, and a transepithelial chloride gradient was applied. Test compounds were added to the apical solution. TMEM16A/CaCC was activated by apical addition of ATP or UTP (100 µM). Isc was measured using a voltage/current clamp system. [2] Whole-Cell Patch Clamp Recording: Recordings were performed at room temperature on TMEM16A-expressing FRT cells or human salivary gland A253 cells. The pipette solution contained 130 mM CsCl, 0.5 mM EGTA, 1 mM MgCl₂, 1 mM Tris-ATP, 10 mM HEPES (pH 7.2), and was supplemented with CaCl₂ to achieve 275 nM free Ca²⁺ for channel activation. The bath solution contained 140 mM N-methyl-D-glucamine-Cl, 1 mM CaCl₂, 1 mM MgCl₂, 10 mM glucose, 10 mM HEPES (pH 7.4). Whole-cell currents were elicited by voltage steps from a holding potential of 0 mV to between -100 mV and +100 mV. Inhibition by compounds was assessed by comparing currents before and after application. [2] Cytoplasmic Calcium Measurement: FRT cells in 96-well plates were loaded with Fluo-4 NW dye. Fluorescence was measured using a plate reader. Cells were pretreated with compound or vehicle for 5 min, followed by addition of calcium agonists (100 µM ATP or 1 µM ionomycin). Changes in fluorescence were monitored to assess if compounds affected calcium signaling. [2] Immunoblot Analysis: Cells were lysed, and proteins were resolved by SDS-PAGE, transferred to a membrane, and probed with a primary antibody against TMEM16A. Detection was performed using a secondary antibody with enhanced chemiluminescence. This method was used to confirm TMEM16A protein expression and the efficacy of siRNA knockdown or IL-4 treatment. [2] RNA Interference (RNAi) Knockdown: T84 cells were transfected with a pool of siRNA targeting TMEM16A or a non-targeting control siRNA using a lipid-based transfection reagent. The transfection was repeated at 24 hours. Functional assays (short-circuit current) and immunoblot analysis were performed 48 hours after the second transfection to assess the effect of TMEM16A knockdown. [2] |
| Animal Protocol |
Male and female New Zealand white rabbits (16–20 weeks old, 2.5–4 kg weight) were humanely killed with a lethal injection of pentobarbitone (i.v.). The most proximal 1.5 cm of the urethra was removed and placed in Krebs’ solution and individual ICC were isolated enzymatically as described previously (Sergeant et al. 2000).[2] Dissected strips of rabbit urethra smooth muscle were stored for subsequent use at −20 °C in RNAlater. Immediately prior to isolation of the RNA, tissue samples were transferred to a 1.5-ml tube, snap frozen in liquid nitrogen, and pulverized to yield a dry powder. RNA was isolated from these samples using the RNeasy mini kit. All samples were DNase treated and the purified RNA was eluted with RNase free water. After determination of the RNA concentration using a nanodrop spectrophotometer, the purified RNA was stored at −80 °C. Prior to cDNA synthesis, RNA was denatured for 5 min at 70 °C and then rapidly cooled on ice. RNA was reverse transcribed using the Superscript VILO cDNA synthesis kit according to the manufacturer’s instructions. Rabbit brain tissue was processed in parallel for use as a control, and cDNA was generated from brain-derived RNA as described above.[2] Real-time quantitative PCR (qPCR) was performed using the SYBR Green PCR Master Mix. TMEM16B and F gene-specific primer sets were designed to span exon-exon boundaries present in the known TMEM16 transcripts. TMEM16A primers were designed from 5′ sequence isolated in our laboratory. β-actin was used in qPCR as an endogenous reference gene for sample normalization, and a β-actin primer set was designed accordingly. [2] |
| References |
[1]. Effects of new-generation TMEM16A inhibitors on calcium-activated chloride currents in rabbit urethral interstitial cells of Cajal. Pflugers Arch. 2017 Nov;469(11):1443-1455. [2]. TMEM16A inhibitors reveal TMEM16A as a minor component of calcium-activated chloride channel conductance in airway and intestinal epithelial cells. J Biol Chem. 2011 Jan 21;286(3):2365-74. |
| Additional Infomation |
TMEM16A (ANO1) functions as a calcium-activated chloride channel (CaCC). We developed pharmacological tools to investigate the contribution of TMEM16A to CaCC conductance in human airway and intestinal epithelial cells. A screen of ∼110,000 compounds revealed four novel chemical classes of small molecule TMEM16A inhibitors that fully blocked TMEM16A chloride current with an IC(50) < 10 μM, without interfering with calcium signaling. Following structure-activity analysis, the most potent inhibitor, an aminophenylthiazole (T16A(inh)-A01), had an IC(50) of ∼1 μM. Two distinct types of inhibitors were identified. Some compounds, such as tannic acid and the arylaminothiophene CaCC(inh)-A01, fully inhibited CaCC current in human bronchial and intestinal cells. Other compounds, including T16A(inh)-A01 and digallic acid, inhibited total CaCC current in these cells poorly, but blocked mainly an initial, agonist-stimulated transient chloride current. TMEM16A RNAi knockdown also inhibited mainly the transient chloride current. In contrast to the airway and intestinal cells, all TMEM16A inhibitors fully blocked CaCC current in salivary gland cells. We conclude that TMEM16A carries nearly all CaCC current in salivary gland epithelium, but is a minor contributor to total CaCC current in airway and intestinal epithelia. The small molecule inhibitors identified here permit pharmacological dissection of TMEM16A/CaCC function and are potential development candidates for drug therapy of hypertension, pain, diarrhea, and excessive mucus production.[2] T16Ainh-A01 was identified through a target-based high-throughput screen of ~110,000 compounds using TMEM16A-transfected cells. [2] It belongs to a chemical class of 2-amino,4-phenylthiazole derivatives linked via a thio-acetyl moiety to a second heterocycle (e.g., pyrimidine). Structure-activity relationship (SAR) analysis indicated that substitutions on the pyrimidine ring at the 3-, 4-, and 5-positions were tolerated, yielding analogs with IC₅₀ values as low as ~1 µM. [2] The study concluded that TMEM16A, inhibited by T16Ainh-A01, is a minor component of the total CaCC conductance in human airway and intestinal epithelial cells, responsible primarily for an initial transient chloride current following agonist stimulation. In contrast, TMEM16A carries nearly all CaCC current in salivary gland epithelial cells. [2] Treatment of airway epithelial cells with IL-4 significantly upregulated TMEM16A expression and increased the T16Ainh-A01-sensitive current component. [2] The findings suggest that direct activators of TMEM16A may not be effective for cystic fibrosis therapy due to its minor role in total airway CaCC conductance, but phenotype-based screens are needed to identify major airway CaCC(s). [2] |
Solubility Data
| Solubility (In Vitro) |
DMSO : ~83.33 mg/mL (~200.06 mM) DMF :≥ 10 mg/mL (~24.01 mM) |
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 2.08 mg/mL (4.99 mM) (saturation unknown) in 10% DMSO + 90% Corn Oil (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 20.8 mg/mL clear DMSO stock solution to 900 μL of corn oil and mix evenly. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.4008 mL | 12.0042 mL | 24.0085 mL | |
| 5 mM | 0.4802 mL | 2.4008 mL | 4.8017 mL | |
| 10 mM | 0.2401 mL | 1.2004 mL | 2.4008 mL |