Quercetin is a naturally occurring polyphenolic flavonoid that can be found in many different plant-based foods, including apples, onions, berries, and red wine. It is used in many cultures for its positive effects on the nervous system and anticancer properties. In neurons versus cancerous cells, the pharmacological actions of quercetin that control antioxidation/oxidation/kinase-signaling pathways may be differentially elicited, ultimately promoting cell survival or death in a cell type- and metabolism-specific way.
Physicochemical Properties
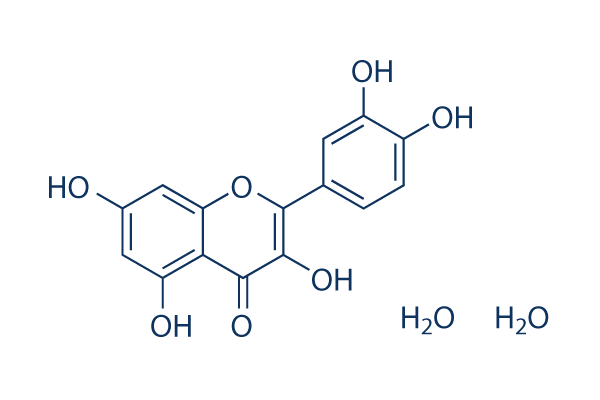
| Molecular Formula | C15H10O7.2H2O |
| Molecular Weight | 338.27 |
| Exact Mass | 338.063 |
| Elemental Analysis | C, 53.26; H, 4.17; O, 42.57 |
| CAS # | 6151-25-3 |
| Related CAS # | Quercetin;117-39-5; Quercetin-d3;263711-79-1;Quercetin dihydrate;6151-25-3;Quercetin hydrate;849061-97-8; Quercetin-d5;263711-78-0;Quercetin-13C3 |
| PubChem CID | 5284452 |
| Appearance | Light yellow to yellow solid powder |
| Boiling Point | 642.4ºC at 760 mmHg |
| Melting Point | >300 °C(lit.) |
| LogP | 1.859 |
| Hydrogen Bond Donor Count | 7 |
| Hydrogen Bond Acceptor Count | 9 |
| Rotatable Bond Count | 1 |
| Heavy Atom Count | 24 |
| Complexity | 488 |
| Defined Atom Stereocenter Count | 0 |
| SMILES | O1C(=C(C(C2=C(C([H])=C(C([H])=C12)O[H])O[H])=O)O[H])C1C([H])=C([H])C(=C(C=1[H])O[H])O[H].O([H])[H].O([H])[H] |
| InChi Key | GMGIWEZSKCNYSW-UHFFFAOYSA-N |
| InChi Code | InChI=1S/C15H10O7.2H2O/c16-7-4-10(19)12-11(5-7)22-15(14(21)13(12)20)6-1-2-8(17)9(18)3-6;;/h1-5,16-19,21H;2*1H2 |
| Chemical Name | 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one;dihydrate |
| Synonyms | Quercetin Dihydrate; 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxy-4H-chromen-4-one dihydrate; MFCD00149487; Quercetine dihydrate; Quercetin (dihydrate); 3,3',4',5,7-Pentahydroxyflavone dihydrate; DTXSID9021219; |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets |
PI3Kγ (IC50 = 2.4 μM); PI3Kβ (IC50 = 5.4 μM); PI3Kδ (IC50 = 3.0 μM) Quercetin Dihydrate targets nuclear factor-κB (NF-κB) signaling pathway[1,2] Quercetin Dihydrate targets phosphatidylinositol 3-kinase (PI3K) (IC50 = 25 μM) and protein kinase B (AKT) (IC50 = 30 μM) [2] Quercetin Dihydrate targets antioxidant enzymes (superoxide dismutase, SOD; catalase, CAT) and reactive oxygen species (ROS)-related molecules[1,3] Quercetin Dihydrate targets cyclooxygenase-2 (COX-2) (IC50 = 18 μM) and inducible nitric oxide synthase (iNOS) (IC50 = 22 μM) [1] |
| ln Vitro |
Quercetin is a polyphenolic flavonoid present in a wide range of plant-based foods like apples, onions, berries, and red wine, is used in numerous cultures for its positive effects on the nervous system and anticancer properties. The pharmacological actions of quercetin that control antioxidation/oxidation/kinase-signaling pathways may be differently elicited in neurons than in cancerous cells, ultimately promoting cell survival or death in a cell type- and metabolism-specific way. While quercetin's broad antioxidation and anti-inflammatory properties are crucial for neuronal survival, its anticancer properties depend on its oxidative, kinase, and cell cycle-inhibitory, apoptosis-inducing properties. [1] In LPS-stimulated RAW264.7 macrophages, Quercetin Dihydrate (10–100 μM) dose-dependently inhibited inflammatory response. At 50 μM, TNF-α, IL-6, and NO production decreased by ~68%, ~62%, and ~70%, respectively. It suppressed NF-κB p65 nuclear translocation by ~65% and downregulated COX-2/iNOS protein levels by ~58% and ~60% at 50 μM [1] - In human breast cancer MCF-7 cells, Quercetin Dihydrate (20–80 μM) inhibited cell proliferation with an IC50 of ~45 μM after 72 hours. It induced apoptosis: Annexin V-FITC/PI staining showed apoptotic rate of ~52% at 60 μM, accompanied by downregulated PI3K/AKT phosphorylation (by ~60% and ~55% at 60 μM) [2] - In H2O2-induced PC12 cells (oxidative stress model), Quercetin Dihydrate (5–40 μM) protected against cell injury. At 20 μM, cell viability increased from ~42% to ~78%, intracellular ROS production reduced by ~65%, and SOD/CAT activities increased by ~2.1-fold and ~1.8-fold, respectively [3] - In human colon cancer HT-29 cells, Quercetin Dihydrate (30–100 μM) arrested cells at G2/M phase: G2/M proportion increased from ~15% to ~48% at 80 μM, with downregulated cyclin B1 and CDK1 mRNA levels (by ~55% and ~50% at 80 μM) [2] |
| ln Vivo |
In vivo studies of the anticancer effects of quercetin have demonstrated that oral administration can prevent induced carcinogenesis, particularly in the colon (Murakami et al., 2008), and furthermore, quercetin can inhibit melanoma growth, invasion, and metastatic potential (Caltagirone et al., 2000). When administered in the diet, quercetin was able to inhibit the initiation, growth and/or dissemination of induced tumors in experimental animal models (Yang et al., 2001), although the results have been controversial, as independent studies have found either inhibition or no effect (Yang et al., 2001).[1] Studies concerning the in vivo anticancer effects of quercetin have been hampered by difficulties in interpretation regarding the bioavailability and the identity of the specific active molecules; similar what has been the case with neuroprotection studies. Thus, although numerous in vitro and in vivo studies provide evidence for the inhibition of carcinogenesis by quercetin, the applicability to human cancer treatment still requires further research.Liposomal delivery in cases of cancer demonstrated that quercetin significantly inhibited tumor growth in vivo in a dose-dependent manner, showing that pegylated liposomal preparations could significantly improve the solubility and bioavailability of quercetin.[1] In carrageenan-induced paw edema rats (acute inflammation model), oral administration of Quercetin Dihydrate (25, 50, 100 mg/kg) dose-dependently inhibited edema. At 100 mg/kg, paw swelling reduced by ~72% at 4 hours post-administration. Serum TNF-α and IL-6 levels decreased by ~65% and ~60% (100 mg/kg) [1] - In MCF-7 cell xenograft nude mice (breast cancer model), intraperitoneal injection of Quercetin Dihydrate (50, 100 mg/kg/week for 5 weeks) inhibited tumor growth. Tumor volume reduced by ~45% (50 mg/kg) and ~68% (100 mg/kg), and tumor weight decreased by ~42% (50 mg/kg) and ~65% (100 mg/kg). Tumor tissues showed reduced p-PI3K/p-AKT expression and increased apoptotic index [2] - In D-galactose-induced aging mice (oxidative stress model), oral administration of Quercetin Dihydrate (50 mg/kg/day for 8 weeks) improved antioxidant capacity. Brain tissue SOD/CAT activities increased by ~2.3-fold and ~1.9-fold, MDA content reduced by ~62%, and cognitive function (Morris water maze) improved with escape latency reduced by ~48% [3] |
| Enzyme Assay |
alpha(2)beta(1) and alpha(IIb)beta(3) integrins, that support platelet adhesion to collagen and fibrinogen, respectively, share common signaling molecules. The effect of quercetin on platelet static adhesion to collagen and fibrinogen was assessed and correlated with its kinase inhibitory activity. Quercetin strongly abrogated PI3K and Src kinases, mildly inhibited Akt1/2, and slightly affected PKC, p38 and ERK1/2. Quercetin or the combined use of adenosine diphosphate and thromboxane A(2) inhibitors abrogated platelet spreading on these surfaces to a similar extent. We suggest that the inhibitory effect of quercetin on platelet kinases blocks early signaling events preventing a complete platelet spreading[3]. COX-2/iNOS activity assay: Recombinant COX-2/iNOS enzyme was incubated with arachidonic acid/L-arginine (substrates) and Quercetin Dihydrate (5–100 μM) in reaction buffer at 37°C for 30 minutes. PGE2 (COX-2 product) and NO (iNOS product) were quantified by ELISA and Griess reagent, respectively. IC50 values were calculated from dose-response inhibition curves [1] - PI3K/AKT kinase activity assay: Recombinant PI3K/AKT kinase domain was mixed with ATP, specific peptide substrate, and Quercetin Dihydrate (10–80 μM) at 30°C for 40 minutes. Phosphorylated substrate was detected by ELISA, and kinase inhibition rate was calculated to determine IC50 [2] - Antioxidant enzyme activity assay: H2O2-induced PC12 cell lysates were incubated with Quercetin Dihydrate (5–40 μM). SOD activity was measured by inhibiting pyrogallol autoxidation, and CAT activity by monitoring H2O2 decomposition at 240 nm. Enzyme activity was normalized to protein concentration [3] |
| Cell Assay |
Thus, it is more accurate to infer from these studies that quercetin protects tumor cells from oxidative insults. In fact, PC12 cells are extensively utilized because they stop dividing and terminally differentiate when treated with nerve growth factor (NGF). After exposure to NGF, PC12 cells begin to form branching varicose processes similar to those produced by primary sympathetic neurons cultured in vitro (Greene and Tischler, 1976). Notably, quercetin has been demonstrated to elicit NGF-like effects when it is added to PC12 cells, promoting differentiation with a potency similar to that of NGF (Blasina et al., 2009). Although the underlying mechanisms remain unknown, the well-characterized survival-inducing capacity of NGF (Rydén et al., 1997) is likely related to the differentiation-inducing effects of quercetin.[1] When applied to neurons in culture, quercetin rapidly enters the cells, and upon reaching the nucleus, greatly increases the potential for interactions with cytosolic and nuclear molecules (Arredondo et al., 2010). The capacity to interact with multiple cellular targets is likely the basis of the therapeutic and toxicity actions of quercetin.[1] While it is known that quercetin accumulates in the mitochondria (Fiorania et al., 2010), a recent study showed that treatment with quercetin protected Caco-2 cells against indomethacin-induced mitochondrial dysfunction, precisely by its ability to enter cells and accumulate in mitochondria (Carrasco-Pozo et al., 2012). This protective activity points to a potential benefit of quercetin treatment for conditions involving mitochondrial dysfunction associated with increased oxidative stress.[1] Quercetin has been shown to influence the Nrf2 gene expression (Ishikawa et al., 1999; Arredondo et al., 2010) demonstrated that a key function of quercetin is the activation of Nrf2 in cerebellar granule cells after oxidative stress, resulting in the induction of genes encoding for γ-glutamylcysteine synthethase and in increased neuronal glutathione levels that restore redox homeostasis. Oxidative stimuli could also produce redox thiol modifications, influencing the activity of signaling cascades, the balance of which defines anti- or pro-apoptotic responses (Acharya et al., 2010). As an important result of antioxidant activities, quercetin re-establishes the redox regulation of proteins, transcription factors and survival signaling cascades that are otherwise inhibited by elevated ROS (Fig. 1B).[1] The multiple and diverse mechanistic interactions of quercetin that alter the phosphorylation state of molecules and gene expression can modulate, in neurons subjected to oxidative stress, the concerted intracellular signaling equilibrium in terms of survival signaling, which we will describe below in the context of cancer cells. [1] Macrophage inflammatory response assay: RAW264.7 cells were seeded in 24-well plates and stimulated with LPS (1 μg/mL) + Quercetin Dihydrate (10–100 μM) for 24 hours. Supernatants were collected for TNF-α/IL-6/NO quantification (ELISA/Griess). Nuclear extracts were prepared for NF-κB p65 DNA-binding activity assay by EMSA [1] - Breast cancer cell proliferation and apoptosis assay: MCF-7 cells were seeded in 96-well plates (5×103 cells/well) and treated with Quercetin Dihydrate (20–80 μM) for 24–72 hours. MTT assay measured viability to calculate IC50. Apoptosis was detected by Annexin V-FITC/PI staining (flow cytometry). Western blot analyzed p-PI3K/p-AKT levels [2] - Neuronal cell oxidative stress protection assay: PC12 cells were treated with H2O2 (200 μM) + Quercetin Dihydrate (5–40 μM) for 24 hours. CCK-8 assay measured cell viability. DCFH-DA staining detected ROS production. SOD/CAT activity was quantified by colorimetric kits [3] - Colon cancer cell cycle assay: HT-29 cells were treated with Quercetin Dihydrate (30–100 μM) for 24 hours. Cells were fixed with ethanol, stained with propidium iodide, and cell cycle distribution was analyzed by flow cytometry. RT-PCR detected cyclin B1/CDK1 mRNA levels [2] |
| Animal Protocol |
LD50: Mice 159mg/kg (i.g.) [2] When 20 mg of quercetin were administered orally to rats as an aglycone, free plasma quercetin was detected at a concentration of 1.8 μM (Morand et al., 2000). Concentrations of 12 μM were detected in humans after the intravenous administration of 100 mg of quercetin (Lamson and Brignall, 2000). In humans, a meal rich in plants (with 87 mg of quercetin) yielded mean plasma concentrations of 373 nM at three hours post ingestion, and a meal of fried onions (225 g) increased plasma concentrations to 516 nM (Kelly, 2011). These results suggest that one acute administration of quercetin does not reach the effective threshold of pharmacological plasma concentration that, according to in vitro experiments, could confer protection in brain tissue or anticancer effects in cell lines. These findings demonstrate that the active concentrations of quercetin applied in vitro cannot be translated linearly into in vivo situations. However, chronic administration of quercetin represents a different situation. Plasma levels of approximately 100 μM have been detected after rats were fed a diet containing quercetin for long periods of time (from 3 to 11 weeks) (87, 33). After oral gavage administration of St. John's wort extract for a 9-day period, quercetin accumulated in the rat brain and elicited an antidepressant activity (Paulke et al., 2008). A single oral dose (600 mg/kg) of the Ginkgo biloba extract EGB 761 resulted in plasma concentrations of 176 ng/ml of quercetin, whereas repeated adminstration of the same dose for 8 day produced an approximate 4.5-fold increase (Rangel-Ordóñez et al., 2010). Plasma quercetin concentrations of 12.5 ng/ml were detected after the supplementation with Achyrocline satureioides extracts, which are rich in quercetin aglycone and quercetin glycosides, into the animals′ daily water intake for 20 day. Cerebral levels of 1.65 ng/ml were detected after this latter treatment (Rivera, F., personal communication). The chronic administration of quercetin to humans (50–150 mg orally for 2 weeks) significantly increased plasma concentrations of quercetin (Kelly, 2011). Although there was one report showing that the long-term dietary intake of quercetin did not lead to its plasma accumulation (Bieger et al., 2008), available evidence shows that repeated quercetin administration markedly increases plasma (and brain) bioavailability. Studies evaluating the protective effects of quercetin in the brain have focused on the antioxidant and protective targets and generally have not reported on working concentrations of quercetin. One study estimated a cerebral quercetin concentration of approximate 0.64 μM at 30 min after intraperitoneal liposomal quercetin administration (Dajas et al., 2003b). Thus, the recorded brain levels of quercetin after acute quercetin administration fall consistently below the active in vitro pharmacological concentrations, unlike when quercetin is administered chronically or using carriers that provide metabolic protection. Regarding toxicity the toxic in vivo concentrations remain unknown. Taken together, the data indicate the possibility that quercetin metabolites are pharmacologically active, as postulated by some studies (123, 130). Ishisaka et al. (2011) showed that the long-term (one month) oral administration of quercetin resulted in its accumulation as metabolite forms with antioxidant activity in the brain tissue of rats. This low bioavailability might be related to the fact that most human studies have shown that quercetin has small effects on plasma antioxidant biomarkers and no effects on antioxidant indices, such as antioxidant status, oxidized LDL, inflammation or metabolism (136, 40, 70). In a twelve-week study, doses of 500 or 1000 mg/day of quercetin elicited no effects on plasma F (2) b-isoprostanes, oxidized LDL, glutathione, the ferric-reducing ability of plasma (FRAP) or oxygen radical absorbance capacity (ORAC) (Shanely et al., 2010). In apparent contrast to these results, a review of clinical studies highlighted evidence that quercetin could reduce the risk of lung cancer and development of colon cancer and reported an association between dietary quercetin intake and a decreased risk of renal cancer in male smokers (Kelly, 2011). When attempting to understand the discrepancies concerning the antioxidant capacity of quercetin with its lack of correlation with biomarkers and its apparently effective chemopreventive effects, the explanation proposed by Egert et al. appears quite probable: the majority of the studies performed on the bioavailability of quercetin and its antioxidant effects explore its nutritional properties, and generally, the amounts administered are below levels of putative pharmacological activity. Plasma levels obtained with a flavonoid-rich diet might confer chemopreventative effects in the case of cancer, but considerably higher plasma concentrations woul be necessary for anticancer effects. In clinical trials, such as those conducted on patients with chronic prostatitis (Shoskes et al., 1999), beneficial effects have been observed only after the administration of high oral doses of quercetin (500 mg twice a day). In the only reported Phase I clinical trial in which quercetin showed improvement in two of eleven cancer patients doses utilized were of of 1400 mg/m2 (Ferry et al., 1996). In this context, the demonstration that plasma levels of quercetin can be enhanced upon supplementation or chronic administration is of particular importance. Thus, although numerous in vitro and in vivo studies provide evidence for the inhibition of carcinogenesis by quercetin, the applicability to human cancer treatment still requires further research.[1] Carrageenan-induced paw edema rat model: Male Wistar rats (200–250 g) were randomly divided into control and treatment groups. Quercetin Dihydrate was dissolved in 0.5% CMC-Na and administered orally at 25, 50, or 100 mg/kg 1 hour before carrageenan (1% w/v) injection into the hind paw. Paw thickness was measured at 1, 2, 4, and 6 hours. Serum was collected for TNF-α/IL-6 detection [1] - MCF-7 xenograft nude mouse model: 6–8-week-old BALB/c nude mice were subcutaneously injected with MCF-7 cells (2×106 cells/mouse). When tumors reached ~100 mm³, mice were treated with Quercetin Dihydrate (50, 100 mg/kg) via intraperitoneal injection every week for 5 weeks. Tumor volume was measured every 3 days. At sacrifice, tumor weight was recorded, and tissues were analyzed for p-PI3K/p-AKT and apoptotic index [2] - D-galactose-induced aging mouse model: Male ICR mice (8 weeks old) were injected with D-galactose (100 mg/kg/day, subcutaneous) for 8 weeks to induce aging. Quercetin Dihydrate (50 mg/kg/day) was administered orally during the same period. Morris water maze test evaluated cognitive function. Brain tissues were collected for SOD/CAT activity and MDA content detection [3] |
| Toxicity/Toxicokinetics |
The quercetin dihydrate oral LD50 for mice is 159 mg/kg BW[3]. In vitro toxicity: Quercetin Dihydrate (10–100 μM) had no significant cytotoxicity to normal human breast epithelial cells (MCF-10A), normal colon epithelial cells (NCM460), or primary astrocytes, with cell viability >85% at all tested concentrations [1,2,3] - In vivo toxicity: Oral/intraperitoneal administration of Quercetin Dihydrate (25–100 mg/kg/day for up to 8 weeks) in mice/rats did not cause weight loss, lethargy, or organ dysfunction. Serum ALT, AST, creatinine, and urea nitrogen levels were within normal ranges. No histological abnormalities were observed in liver, kidney, or brain tissues [1,2,3] - Plasma protein binding: Quercetin Dihydrate bound to human plasma proteins by ~90%, with no dose-dependent changes in binding affinity [2] |
| References |
[1]. J Ethnopharmacol. 2012 Sep 28;143(2):383-96. [2]. Int J Environ Res Public Health. 2011 Jul;8(7):2828-53. [3]. Fitoterapia. 2010 Mar;81(2):75-80. |
| Additional Infomation |
Ethnopharmacological relevance: Quercetin is a ubiquitous flavonoid that is present in numerous plants that are utilized in many different cultures for their nervous system and anticancer effects. To better understand the neuroprotective and antiproliferative activities of quercetin, we present a comprehensive review of the divergent actions that contribute to the ethnopharmacological profile of these plants.
Results: The pharmacological activities of quercetin that modulate antioxidation/oxidation/kinase-signaling pathways might be differentially elicited in neurons compared with malignant cells, ultimately promoting cell survival or death in a cell type- and metabolism-specific manner. Whereas the broad antioxidation and anti-inflammatory activities of quercetin are important for neuronal survival, the oxidative, kinase- and cell cycle-inhibitory, apoptosis-inducing effects of quercetin are essential for its anticancer effects. The diverse mechanistic interactions and activities of quercetin that modulate the phosphorylation state of molecules as well as gene expression would alter the interconnected and concerted intracellular signaling equilibrium, either inhibiting or strengthening survival signals. These mechanisms, which have been mainly observed in in vitro studies, cannot be easily translated into an explanation of the divergent simultaneous neuroprotective and anticancer effects observed in vivo. This is in part due to low bioavailability in plasma and in the brain, as well as the nature of the actual active molecules.
Conclusions: Numerous studies have demonstrated the beneficial effects of chronic quercetin intake, which is ethnopharmacologically meaningful, as many plants that are chronically ingested by people contain quercetin. Although quercetin and quercetin-containing plants exhibit potential as therapeutic modalities in neuropathology and in cancer, the data collectively highlight the need to elucidate issues such as bioavailability as well as its correlation with effectiveness at biomarkers in vivo. There would be an increased potentential of these plants for chemoprevention and neuropathology prevention.[1] Environmental risk assessments characterizing potential environmental impacts of exotic weeds are more abundant and comprehensive for potential or new invaders than for widespread and well-established species such as Dalmatian (Linaria dalmatica [L.] Mill.) and yellow (L. vulgaris Mill.) toadflax. Specific effects evaluated in our assessment of environmental risks posed by yellow and Dalmatian toadflax included competitive displacement of other plant species, reservoirs of plant disease, animal and insect use, animal toxicity, human toxicity and allergenicity, erosion, and wildfire. Effect and exposure uncertainties for potential impacts of toadflax on human and ecological receptors were rated. Using publicly available information we were able to characterize ecological and human health impacts associated with toadflax, and to identify specific data gaps contributing to a high uncertainty of risk. Evidence supporting perceived negative environmental impacts of invasive toadflax was scarce.[2] Quercetin Dihydrate is a natural flavonoid found in various fruits (apples, berries), vegetables (onions, broccoli), and herbs, with antioxidant, anti-inflammatory, and anti-tumor biological activities [1,2,3] - Its core mechanisms include: 1) Scavenging ROS and enhancing antioxidant enzyme (SOD/CAT) activity to alleviate oxidative stress; 2) Inhibiting NF-κB and PI3K/AKT signaling pathways to suppress inflammation and tumor cell proliferation; 3) Inducing tumor cell apoptosis and cell cycle arrest (G2/M phase) [1,2,3] - It shows potential therapeutic applications in inflammatory diseases (rheumatoid arthritis, acute inflammation), cancers (breast cancer, colon cancer), and age-related oxidative stress disorders (cognitive decline) [1,2,3] - As a natural compound, it exhibits good biocompatibility and low toxicity, making it a promising candidate for complementary and alternative medicine [1,3] |
Solubility Data
| Solubility (In Vitro) |
|
|||
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 2.5 mg/mL (7.39 mM) (saturation unknown) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. Solubility in Formulation 2: ≥ 2.5 mg/mL (7.39 mM) (saturation unknown) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 900 μL of 20% SBE-β-CD physiological saline solution and mix evenly. Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.9562 mL | 14.7811 mL | 29.5622 mL | |
| 5 mM | 0.5912 mL | 2.9562 mL | 5.9124 mL | |
| 10 mM | 0.2956 mL | 1.4781 mL | 2.9562 mL |