Physicochemical Properties
| Molecular Formula | C30H33N3O5S |
| Molecular Weight | 547.67 |
| Exact Mass | 547.214 |
| CAS # | 925701-46-8 |
| PubChem CID | 15953870 |
| Appearance | Light yellow to khaki solid powder |
| Density | 1.3±0.1 g/cm3 |
| Boiling Point | 786.6±60.0 °C at 760 mmHg |
| Flash Point | 429.5±32.9 °C |
| Vapour Pressure | 0.0±2.7 mmHg at 25°C |
| Index of Refraction | 1.645 |
| LogP | 5.9 |
| Hydrogen Bond Donor Count | 1 |
| Hydrogen Bond Acceptor Count | 8 |
| Rotatable Bond Count | 5 |
| Heavy Atom Count | 39 |
| Complexity | 972 |
| Defined Atom Stereocenter Count | 2 |
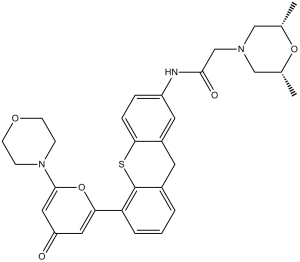
| SMILES | C[C@@H]1CN(C[C@@H](O1)C)CC(=O)NC2=CC3=C(C=C2)SC4=C(C3)C=CC=C4C5=CC(=O)C=C(O5)N6CCOCC6 |
| InChi Key | SCELLOWTHJGVIC-BGYRXZFFSA-N |
| InChi Code | InChI=1S/C30H33N3O5S/c1-19-16-32(17-20(2)37-19)18-28(35)31-23-6-7-27-22(13-23)12-21-4-3-5-25(30(21)39-27)26-14-24(34)15-29(38-26)33-8-10-36-11-9-33/h3-7,13-15,19-20H,8-12,16-18H2,1-2H3,(H,31,35)/t19-,20+ |
| Chemical Name | 2-[(2S,6R)-2,6-dimethylmorpholin-4-yl]-N-[5-(6-morpholin-4-yl-4-oxopyran-2-yl)-9H-thioxanthen-2-yl]acetamide |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets | ATM 6.3 nM (IC50) DNA-PKcs 1.7 μM (IC50) |
| ln Vitro | KU-60019 is KU-55933's enhanced counterpart. KU-55933 is highly selective for ATM kinase, with an in vitro IC50 of 13 nM and a Ki of 2.2 nM, utilizing a panel of 60 protein kinases. An enhanced ATM kinase inhibitor, KU-60019 has an IC50 of 6.3 nM, which is roughly half that of KU-55933. DNA-PKcs and ATR had IC50 values of 1.7 and >10 μM, respectively, over 270 and 1600 times greater than ATM. When it comes to preventing radiation-induced phosphorylation of important ATM targets in human glioma cells, KU-60019 is ten times more efficient than KU-55933. When exposed to 10 μM of KU-55933, human U87 glioma cells showed complete inhibition of p53 (S15) phosphorylation, but not at 3 μM. In contrast, 1 hour after irradiation, γ-H2AX levels were only slightly reduced. On the other hand, at 3 μM and 1 μM, respectively, KU-60019 completely and partially blocked p53 phosphorylation [1]. |
| ln Vivo | PTEN-deficient control tumors grew four times larger than PTEN wild-type controls, however when treated with KU-60019, PTEN-deficient tumor development slowed statistically significantly. This reduction of growth was most noticeable during the first few days of the experiment (days 5–12) and following the administration of KU–60019 (days 1–5) [2]. |
| References |
[1]. Improved ATM kinase inhibitor KU-60019 radiosensitizes glioma cells, compromises insulin, AKT and ERK prosurvival signaling, and inhibits migration and invasion. Mol Cancer Ther. 2009 Oct;8(10):2894-902. [2]. Mechanistic Rationale to Target PTEN-Deficient Tumor Cells with Inhibitors of the DNA Damage Response Kinase ATM. Cancer Res. 2015 Jun 1;75(11):2159-65. |
Solubility Data
| Solubility (In Vitro) |
DMSO: 100 mg/mL (182.59 mM) Ethanol: 10 mg/mL (18.26 mM) |
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 2.5 mg/mL (4.56 mM) (saturation unknown) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. Solubility in Formulation 2: ≥ 2.5 mg/mL (4.56 mM) (saturation unknown) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 900 μL of 20% SBE-β-CD physiological saline solution and mix evenly. Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.8259 mL | 9.1296 mL | 18.2592 mL | |
| 5 mM | 0.3652 mL | 1.8259 mL | 3.6518 mL | |
| 10 mM | 0.1826 mL | 0.9130 mL | 1.8259 mL |