Physicochemical Properties
| Molecular Formula | C18H18N4O2S3 |
| Molecular Weight | 418.548 |
| Exact Mass | 418.059 |
| Elemental Analysis | C, 51.65; H, 4.33; N, 13.39; O, 7.65; S, 22.98 |
| CAS # | 688353-45-9 |
| PubChem CID | 3244448 |
| Appearance | White to off-white solid powder |
| Density | 1.6±0.1 g/cm3 |
| Index of Refraction | 1.783 |
| LogP | 3.99 |
| Hydrogen Bond Donor Count | 1 |
| Hydrogen Bond Acceptor Count | 7 |
| Rotatable Bond Count | 4 |
| Heavy Atom Count | 27 |
| Complexity | 711 |
| Defined Atom Stereocenter Count | 0 |
| InChi Key | RBFDSBJDWZOTGR-UHFFFAOYSA-N |
| InChi Code | InChI=1S/C18H18N4O2S3/c1-9-4-5-11-13(6-9)27-17(19-11)21-14(23)8-25-18-20-12-7-10(2)26-15(12)16(24)22(18)3/h4-6,10H,7-8H2,1-3H3,(H,19,21,23) |
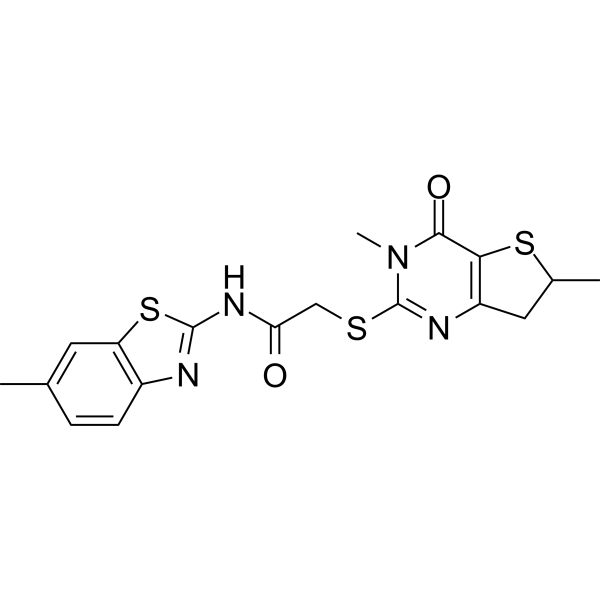
| Chemical Name | 2-[(3,6-dimethyl-4-oxo-6,7-dihydrothieno[3,2-d]pyrimidin-2-yl)sulfanyl]-N-(6-methyl-1,3-benzothiazol-2-yl)acetamide |
| Synonyms | IWP-12; IWP 12; 688353-45-9; IWP12; IWP-12; MLS000091043; SMR000025621; 2-[(3,6-dimethyl-4-oxo-6,7-dihydrothieno[3,2-d]pyrimidin-2-yl)sulfanyl]-N-(6-methyl-1,3-benzothiazol-2-yl)acetamide; IWP 12; N-(6-Methyl-2-benzothiazolyl)-2-[(3,4,6,7-tetrahydro-3,6-dimethyl-4-oxothieno[3,2-d]pyrimidin-2-yl)thio]acetamide; IWP12 |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets | PORCN (porcupine); WNT |
| ln Vitro | Aberrant activation of Wnt/β-catenin signaling plays a key role in the onset and development of hepatocellular carcinomas (HCC), with about half of them acquiring mutations in either CTNNB1 or AXIN1. However, it remains unclear whether these mutations impose sufficient β-catenin signaling or require upstream Wnt ligand activation for sustaining optimal growth, as previously suggested for colorectal cancers. Using a panel of nine HCC cell lines, we show that siRNA-mediated knockdown of β-catenin impairs growth of all these lines. Blocking Wnt secretion, by either treatment with the IWP-12 porcupine inhibitor or knockdown of WLS, reduces growth of most of the lines. Unexpectedly, interfering with Wnt secretion does not clearly affect the level of β-catenin signaling in the majority of lines, suggesting that other mechanisms underlie the growth-suppressive effect. However, IWP-12 treatment did not induce autophagy or endoplasmic reticulum (ER) stress, which may have resulted from the accumulation of Wnt ligands within the ER. Similar results were observed for colorectal cancer cell lines used for comparison in various assays. These results suggest that most colorectal and liver cancers with mutations in components of the β-catenin degradation complex do not strongly rely on extracellular Wnt ligand exposure to support optimal growth. In addition, our results also suggest that blocking Wnt secretion may aid in tumor suppression through alternative routes currently unappreciated. |
| ln Vivo | Despite its modest metabolic stability in mouse-derived plasma, IWP-L6 (27) was highly active in zebrafish. We had previously shown that both the tankyrase (Tnks) inhibitor IWR-1 and the Porcn inhibitor IWP-12 (7) effectively block the regeneration of the tailfin, a Wnt-dependent process, in adult and juvenile fish.3,8,16 We herein show that IWP-L6 (27) exhibited more potent activity (Figure 6). We further show that IWP-L6 (27) and 35, but not 30 and 32, effectively inhibited posterior axis formation, a Wnt/β-catenin dependent developmental process, at low micromolar concentrations (Figure 7). IWP-L6 (27) and 35 are therefore at least 10 times more potent than IWP-12 (7) and 2.5 times more potent than IWR-1 in this in vivo assay.16 While there is only 69% sequence identity between mouse Porcn and zebrafish Porcn, the in vitro EC50 values (Figure 3) measured in mouse fibroblasts (L cells) correlate with the in vivo activity observed in fish but not linearly[1]. |
| Enzyme Assay |
S9 Metabolism Assay. [1] Male ICR/CD-1 mouse S9 fractions were used. 50 μL (1 mg) of S9 protein was added to a 15 mL glass screw cap tube. 700 μL of a 50 mM Tris, pH 7.5 solution, containing the compound of interest was added on ice. The final concentration of compound after addition of all reagents was 2 μM. 250 μL of an NADPH-regenerating system (1.7 mg/mL NADP, 7.8 mg/mL glucose-6-phosphate, 6 U/mL glucose-6-phosphate dehydrogenase in 2% w/v NaHCO3/10 mM MgCl2) was added and the tube placed in a 37 °C shaking water bath. At varying time points after addition of phase I cofactors, the reaction was stopped by the addition of 1 mL of methanol containing an internal standard compound, n-benzylbenzamide, and formic acid. The samples were incubated 10 min at room temperature and then spun once at 975× g in the glass tube. The supernatant was spun a second time at 16,000× g for 5 min in a microcentrifuge. The supernatant was analyzed by LC-MS/MS. Analytical methods were developed for each compound using an AB SCIEX 3200-QTrap, a combination triple quadrupole/ion trap instrument. Compound transitions utilized for quantitation were as follows: IWP-L6 (27): 473.2 to 303.1; aniline: 171.2 to 127.0; n-benzylbenzamide (internal standard): 212.1 to 91.1. A Shimadzu Prominence LC with Agilent C18 XDB column (5 micron, 50 × 4.6 mm) was used for chromatography. The peak areas for IWP-L6 and the aniline were normalized to the n-benzylbenzamide peak area and then the relative amount of compound present at each time point was normalized to the amount present at time 0 and presented as a percentage. Plasma Metabolism Assay. [1] Murine, rat, and human plasma, collected using acidified citrate dextrose (ACD) anticoagulant, were purchased from Bioreclamation (Westbury, NY). IWP-L6 (27) was added to 1 mL of each plasma at a final concentration of 2 μM, the material was aliquoted into eppendorf tubes, and then incubated for the indicated time points in a 37 °C water bath. At each time point, an aliquot was removed and protein precipitated and compound extracted and analyzed as described above. |
| Cell Assay |
MTT Assay[2] A total of 10 mM 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was added to cells seeded in 96-well plates and incubated at 37°C with 5% CO2 for 3 hours. The medium was removed, and 100 μl DMSO was added to each well. The absorbance of each well was read on a microplate absorbance reader at wavelength of 490 nm. For siRNA-mediated knockdown of genes, four independent wells were assayed for each cell line at least two times, whereas for IWP-12 treatment, six independent wells were used. The mean and standard error were calculated for each condition. |
| References |
[1]. The development of highly potent inhibitors for porcupine. J Med Chem. 2013 Mar 28;56(6):2700-4. [2]. Blocking Wnt Secretion Reduces Growth of Hepatocellular Carcinoma Cell Lines Mostly Independent of β-Catenin Signaling. Neoplasia. 2016 Dec;18(12):711-723. |
| Additional Infomation | Porcupine is a member of the membrane-bound O-acyltransferase family of proteins. It catalyzes the palmitoylation of Wnt proteins, a process required for their secretion and activity. We recently disclosed a class of small molecules (IWPs) as the first reported Porcn inhibitors. We now describe the structure-activity relationship studies and the identification of subnanomolar inhibitors. We also report herein the effects of IWPs on Wnt-dependent developmental processes, including zebrafish posterior axis formation and kidney tubule formation.[1] |
Solubility Data
| Solubility (In Vitro) | DMSO : ~25 mg/mL (~59.73 mM) |
| Solubility (In Vivo) |
Solubility in Formulation 1: 2.5 mg/mL (5.97 mM) in 10% DMSO + 40% PEG300 +5% Tween-80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), suspension solution; with sonication. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 25.0 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 + to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.3892 mL | 11.9460 mL | 23.8920 mL | |
| 5 mM | 0.4778 mL | 2.3892 mL | 4.7784 mL | |
| 10 mM | 0.2389 mL | 1.1946 mL | 2.3892 mL |