Physicochemical Properties
| Molecular Formula | C50H46CL2F2N2O23 |
| Molecular Weight | 1151.80266141891 |
| Exact Mass | 1150.183 |
| CAS # | 348079-13-0 |
| PubChem CID | 3626995 |
| Appearance | Typically exists as solid at room temperature |
| Density | 1.5±0.1 g/cm3 |
| Boiling Point | 1087.3±65.0 °C at 760 mmHg |
| Flash Point | 611.4±34.3 °C |
| Vapour Pressure | 0.0±0.3 mmHg at 25°C |
| Index of Refraction | 1.611 |
| LogP | 4.94 |
| Hydrogen Bond Donor Count | 0 |
| Hydrogen Bond Acceptor Count | 27 |
| Rotatable Bond Count | 36 |
| Heavy Atom Count | 79 |
| Complexity | 2270 |
| Defined Atom Stereocenter Count | 0 |
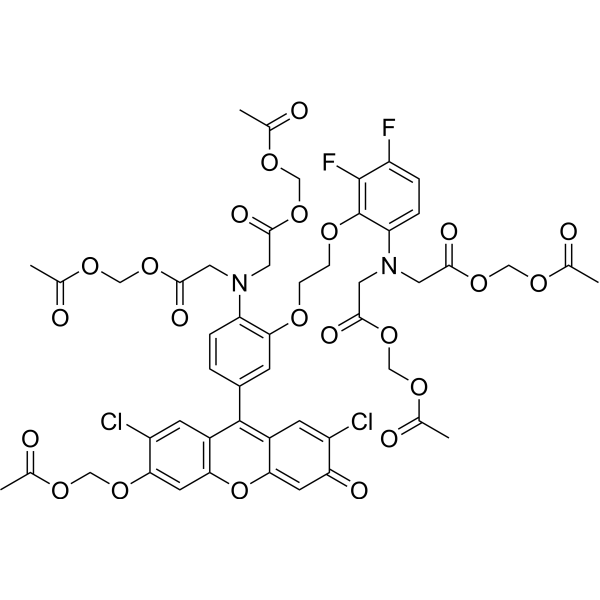
| SMILES | ClC1C(=CC2=C(C=1)C(=C1C=C(C(C=C1O2)=O)Cl)C1C=CC(=C(C=1)OCCOC1C(=C(C=CC=1N(CC(=O)OCOC(C)=O)CC(=O)OCOC(C)=O)F)F)N(CC(=O)OCOC(C)=O)CC(=O)OCOC(C)=O)OCOC(C)=O |
| InChi Key | ABZBPPQDKRLSCZ-UHFFFAOYSA-N |
| InChi Code | InChI=1S/C50H46Cl2F2N2O23/c1-26(57)69-21-74-42-16-41-33(14-35(42)52)48(32-13-34(51)39(62)15-40(32)79-41)31-6-8-37(55(17-44(63)75-22-70-27(2)58)18-45(64)76-23-71-28(3)59)43(12-31)67-10-11-68-50-38(9-7-36(53)49(50)54)56(19-46(65)77-24-72-29(4)60)20-47(66)78-25-73-30(5)61/h6-9,12-16H,10-11,17-25H2,1-5H3 |
| Chemical Name | acetyloxymethyl 2-[4-[3-(acetyloxymethoxy)-2,7-dichloro-6-oxoxanthen-9-yl]-N-[2-(acetyloxymethoxy)-2-oxoethyl]-2-[2-[6-[bis[2-(acetyloxymethoxy)-2-oxoethyl]amino]-2,3-difluorophenoxy]ethoxy]anilino]acetate |
| Synonyms | Fluo-3FF AM; 348079-13-0; acetyloxymethyl 2-[4-[3-(acetyloxymethoxy)-2,7-dichloro-6-oxoxanthen-9-yl]-N-[2-(acetyloxymethoxy)-2-oxoethyl]-2-[2-[6-[bis[2-(acetyloxymethoxy)-2-oxoethyl]amino]-2,3-difluorophenoxy]ethoxy]anilino]acetate; N-[4-[6-[(acetyloxy)methoxy]-2,7-dichloro-3-oxo-3H-xanthen-9-yl]-2-[2-[6-[bis[2-[(acetyloxy)methoxy]-2-oxoethyl]amino]-2,3-difluorophenoxy]ethoxy]phenyl]-N-[2-[(acetyloxy)methoxy]-2-oxoethyl]-glycine(acetyloxy)methylester; HY-D1755; N-[4-[6-[(acetyloxy)methoxy]-2,7-dichloro-3-oxo-3H-xanthen-9-yl]-2-[2-[6-[bis[2-[(acetyloxy)methoxy]-2-oxoethyl]amino]-2,3-difluorophenoxy]ethoxy]phenyl]-N-[2-[(acetyloxy)methoxy]-2-oxoethyl]-glycine(acetyloxy)methyl ester; PD044414; |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets | Fluorecent dye/Ca2+ chelator |
| ln Vitro |
1. Preparation of Fluo-3FF AM Working Solution 1.1 Stock Solution Preparation Dissolve Fluo-3FF AM in 0.025% (w/v) Pluronic F-127/DMSO solution to prepare a 1 mM stock solution. Note: The prepared stock solution should be aliquoted and stored at -20°C or -80°C in the dark. 1.2 Working Solution Preparation Dilute the stock solution with balanced buffer solution to prepare a 5 μM working solution. Note: The working solution concentration can be adjusted according to experimental requirements and should be prepared fresh before use. 2. Cell Staining Procedure 2.1 Culture adherent cells on sterile coverslips in advance. 2.2 During the experiment, remove the coverslips and carefully aspirate the residual medium. 2.3 Add 100 μL of dye working solution, gently swirl to ensure complete coverage of the cell layer, then incubate for 60 minutes. 2.4 After incubation, remove the dye solution and wash the cells with 4°C pre-cooled PSS solution for 60 minutes. |
| Enzyme Assay |
Visualisation of intracellular calcium stores: To visualise the distribution of intracellular calcium stores within the myocytes, the low-affinity (Kd=42 μM) fluorescent Ca2+-sensitive indicator fluo-3FF (Abs/Em=462 nm/526 nm) was used. This dye was selected among other low-affinity Ca2+-sensitive indicators because of its insensitivity to Mg2+ and relatively high photostability. The myocytes were loaded with fluo-3FF by exposure to 5 μM Fluo-3FF AM (diluted from a stock containing 1 mM Fluo-3FF AM and 0.025% (w/v) pluronic F-127 in dimethyl sulphoxide) for 60–90 min at room temperature, followed by 60-min wash in PSS at 4 °C. To visualise three-dimensional (3-D) distribution of the intracellular calcium stores within the cell, z-sectioning protocol (series of x–y images taken at defined intervals along the z-axis) was applied to the myocytes preloaded with the low-affinity Ca2+-sensitive indicator Fluo-3FF (see above). This protocol was comprised of 30–45 individual x–y images each taken from a confocal optical section below 0.8 μm with a z-step of 0.4 μm. Fluo-3FF fluorescence was excited by the 488 nm line of a 200 mW argon ion laser and the emitted fluorescence was detected at wavelengths above 505 nm [1]. |
| Cell Assay |
Intracellular calcium stores of human uterine myocytes in primary and second passage cell culture were visualized using the low-affinity calcium-sensitive fluorescent dye, fluo-3FF. The calcium stores appeared as numerous small (0.2-0.5 microm diameter) focal fluorescences. The stores were not depleted by exposing the cells to oxytocin or ryanodine under standard conditions. The stores were rapidly depleted by oxytocin or ryanodine exposure when sarcoplasmic reticulum (SR) calcium re-uptake was inhibited by pretreatment with thapsigargin. Immunofluorescence experiments indicated that both ryanodine and inositol 1,4,5-trisphosphate (IP(3)) receptors were smoothly distributed throughout the SR, and neither receptor co-localized with the calcium stores. Since IP(3) and ryanodine calcium channels are tightly associated with their receptor, these results suggest that SR calcium release occurs via second messenger channels that are remote from the SR calcium stores. These observations are consistent only with a mechanism for release of calcium stores where the SR serves three functions: (1) as site of calcium storage, (2) as the structure that contains the IP(3)- and ryanodine receptors and their associated release channels, and (3) as a conduit between the calcium stores and the release channels. [2] The dynamics of carbachol (CCh)-induced [Ca(2+)](i) changes was related to the kinetics of muscarinic cationic current (mI(cat)) and the effect of Ca(2+) release through ryanodine receptors (RyRs) and inositol 1,4,5-trisphosphate receptors (IP(3)Rs) on mI(cat) was evaluated by fast x-y or line-scan confocal imaging of [Ca(2+)](i) combined with simultaneous recording of mI(cat) under whole-cell voltage clamp. When myocytes freshly isolated from the longitudinal layer of the guinea-pig ileum were loaded with the Ca(2+)-sensitive indicator fluo-3, x-y confocal imaging revealed CCh (10 microM)-induced Ca(2+) waves, which propagated from the cell ends towards the myocyte centre at 45.9 +/- 8.8 microms(-1) (n = 13). Initiation of the Ca(2+) wave preceded the appearance of any measurable mI(cat) by 229 +/- 55 ms (n = 7). Furthermore, CCh-induced [Ca(2+)](i) transients peaked 1.22 +/- 0.11s (n = 17) before mI(cat) reached peak amplitude. At -50 mV, spontaneous release of Ca(2+) through RyRs, resulting in Ca(2+) sparks, had no effect on CCh-induced mI(cat) but activated BK channels leading to spontaneous transient outward currents (STOCs). In addition, Ca(2+) release through RyRs induced by brief application of 5 mM caffeine was initiated at the cell centre but did not augment mI(cat) (n = 14). This was not due to an inhibitory effect of caffeine on muscarinic cationic channels (since application of 5 mM caffeine did not inhibit mI(cat) when [Ca(2+)](i) was strongly buffered with Ca(2+)/BAPTA buffer) nor was it due to an effect of caffeine on other mechanisms possibly involved in the regulation of Ca(2+) sensitivity of muscarinic cationic channels (since in the presence of 5 mM caffeine, photorelease of Ca(2+) upon cell dialysis with 5 mM NP-EGTA/3.8 mM Ca(2+) potentiated mI(cat) in the same way as in control). In contrast, IP(3)R-mediated Ca(2+) release upon flash photolysis of "caged" IP(3) (30 microM in the pipette solution) augmented mI(cat) (n = 15), even though [Ca(2+)](i) did not reach the level required for potentiation of mI(cat) during photorelease of Ca(2+) (n = 10). Intracellular calcium stores were visualised by loading of the myocytes with the low-affinity Ca(2+) indicator Fluo-3FF AM and consisted of a superficial sarcoplasmic reticulum (SR) network and some perinuclear formation, which appeared to be continuous with the superficial SR. Immunostaining of the myocytes with antibodies to IP(3)R type 1 and to RyRs revealed that IP(3)Rs are predominant in the superficial SR while RyRs are confined to the central region of the cell. These results suggest that IP(3)R-mediated Ca(2+) release plays a central role in the modulation of mI(cat) in the guinea-pig ileum and that IP(3) may sensitise the regulatory mechanisms of the muscarinic cationic channels gating to Ca(2+) [1]. |
| References |
[1]. Regulation of muscarinic cationic current in myocytes from guinea-pig ileum by intracellular Ca2+ release: a central role of inositol 1,4,5-trisphosphate receptors. Cell Calcium. 2004 Nov;36(5):367-86. [2]. Focal sarcoplasmic reticulum calcium stores and diffuse inositol 1,4,5-trisphosphate and ryanodine receptors in human myometrium. Cell Calcium. 1999 Jul-Aug;26(1-2):69-75. |
| Additional Infomation | Tight coupling between IP3-induced Ca2+ release (IICR) and modulation of the muscarinic cationic channel gating suggests sub-plasmalemmal location of SR elements in ileal myocytes. To visualise the spatial arrangements of intracellular calcium stores in living SMCs freshly isolated from the longitudinal layer of the guinea-pig ileum we used the low-affinity Ca2+-sensitive indicator fluo-3FF, which was loaded into the cells by 60–90-min incubation of the myocytes with 5 μM Fluo-3FF AM followed by 60-min wash in PSS to allow time for de-esterification (see Section 2.3). Due to low-affinity of this dye for Ca2+ (Kd=42 μM), the fluorescence signal from regions of the myocytes where [Ca2+]i<1 μM was weak and was eliminated from the images by thresholding, and therefore cytoplasmic Ca2+ was not imaged. To visualise 3-D distribution of the intracellular calcium store elements, a z-sectioning protocol was used (see Section 2.5). It was comprised of 35–40 x–y fluorescence images taken from the confocal optical slice (<0.8 μm) of the myocyte with a z-step (displacement of the objective) of 0.4 μm (n=12). This experimental approach revealed that intracellular calcium stores in ileal myocytes consisted of a well-developed sub-plasmalemmal SR network and some perinuclear formation (probably nuclear envelope and Golgi apparatus), which appeared to be continuous with superficial SR (Fig. 8A). The distribution of the calcium store elements within ileal myocytes was generally similar to that we have recently demonstrated in the rabbit portal vein myocytes using DiOC6 and BODIPY TR-X ryanodine. This pattern of intracellular calcium store arrangement was further confirmed in 35 ileal myocytes by x–y imaging fluo-3FF fluorescence through the middle (confocal optical slice <0.8 μm) of the cell, as shown in Fig. 8B. This data suggest that the SR in ileal myocytes is well developed at a sub-plasmalemmal location and may serve to deliver Ca2+ ions with high precision, both in time and space, to modulate Ca2+-dependent ion channels in the plasma membrane [1]. |
Solubility Data
| Solubility (In Vitro) | May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples |
| Solubility (In Vivo) |
Note: Listed below are some common formulations that may be used to formulate products with low water solubility (e.g. < 1 mg/mL), you may test these formulations using a minute amount of products to avoid loss of samples. Injection Formulations (e.g. IP/IV/IM/SC) Injection Formulation 1: DMSO : Tween 80: Saline = 10 : 5 : 85 (i.e. 100 μL DMSO stock solution → 50 μL Tween 80 → 850 μL Saline) *Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH ₂ O to obtain a clear solution. Injection Formulation 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (i.e. 100 μL DMSO → 400 μLPEG300 → 50 μL Tween 80 → 450 μL Saline) Injection Formulation 3: DMSO : Corn oil = 10 : 90 (i.e. 100 μL DMSO → 900 μL Corn oil) Example: Take the Injection Formulation 3 (DMSO : Corn oil = 10 : 90) as an example, if 1 mL of 2.5 mg/mL working solution is to be prepared, you can take 100 μL 25 mg/mL DMSO stock solution and add to 900 μL corn oil, mix well to obtain a clear or suspension solution (2.5 mg/mL, ready for use in animals). Injection Formulation 4: DMSO : 20% SBE-β-CD in saline = 10 : 90 [i.e. 100 μL DMSO → 900 μL (20% SBE-β-CD in saline)] *Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. Injection Formulation 5: 2-Hydroxypropyl-β-cyclodextrin : Saline = 50 : 50 (i.e. 500 μL 2-Hydroxypropyl-β-cyclodextrin → 500 μL Saline) Injection Formulation 6: DMSO : PEG300 : castor oil : Saline = 5 : 10 : 20 : 65 (i.e. 50 μL DMSO → 100 μLPEG300 → 200 μL castor oil → 650 μL Saline) Injection Formulation 7: Ethanol : Cremophor : Saline = 10: 10 : 80 (i.e. 100 μL Ethanol → 100 μL Cremophor → 800 μL Saline) Injection Formulation 8: Dissolve in Cremophor/Ethanol (50 : 50), then diluted by Saline Injection Formulation 9: EtOH : Corn oil = 10 : 90 (i.e. 100 μL EtOH → 900 μL Corn oil) Injection Formulation 10: EtOH : PEG300:Tween 80 : Saline = 10 : 40 : 5 : 45 (i.e. 100 μL EtOH → 400 μLPEG300 → 50 μL Tween 80 → 450 μL Saline) Oral Formulations Oral Formulation 1: Suspend in 0.5% CMC Na (carboxymethylcellulose sodium) Oral Formulation 2: Suspend in 0.5% Carboxymethyl cellulose Example: Take the Oral Formulation 1 (Suspend in 0.5% CMC Na) as an example, if 100 mL of 2.5 mg/mL working solution is to be prepared, you can first prepare 0.5% CMC Na solution by measuring 0.5 g CMC Na and dissolve it in 100 mL ddH2O to obtain a clear solution; then add 250 mg of the product to 100 mL 0.5% CMC Na solution, to make the suspension solution (2.5 mg/mL, ready for use in animals). Oral Formulation 3: Dissolved in PEG400 Oral Formulation 4: Suspend in 0.2% Carboxymethyl cellulose Oral Formulation 5: Dissolve in 0.25% Tween 80 and 0.5% Carboxymethyl cellulose Oral Formulation 6: Mixing with food powders Note: Please be aware that the above formulations are for reference only. InvivoChem strongly recommends customers to read literature methods/protocols carefully before determining which formulation you should use for in vivo studies, as different compounds have different solubility properties and have to be formulated differently. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 0.8682 mL | 4.3410 mL | 8.6821 mL | |
| 5 mM | 0.1736 mL | 0.8682 mL | 1.7364 mL | |
| 10 mM | 0.0868 mL | 0.4341 mL | 0.8682 mL |