Physicochemical Properties
| Molecular Formula | C22H24N2O10 |
| Molecular Weight | 476.4334 |
| Exact Mass | 476.143 |
| Elemental Analysis | C, 55.46; H, 5.08; N, 5.88; O, 33.58 |
| CAS # | 85233-19-8 |
| Related CAS # | BAPTA tetrasodium;126824-24-6;BAPTA tetrapotassium;73630-08-7; BAPTA;85233-19-8;BAPTA tetrasodium;126824-24-6; 480436-84-8 (cesium) |
| PubChem CID | 104751 |
| Appearance | Off-white to light brown solid powder |
| Density | 1.5±0.1 g/cm3 |
| Boiling Point | 766.6±60.0 °C at 760 mmHg |
| Melting Point | 177-179 °C |
| Flash Point | 417.4±32.9 °C |
| Vapour Pressure | 0.0±2.7 mmHg at 25°C |
| Index of Refraction | 1.657 |
| LogP | 0.13 |
| Hydrogen Bond Donor Count | 4 |
| Hydrogen Bond Acceptor Count | 12 |
| Rotatable Bond Count | 15 |
| Heavy Atom Count | 34 |
| Complexity | 613 |
| Defined Atom Stereocenter Count | 0 |
| InChi Key | FTEDXVNDVHYDQW-UHFFFAOYSA-N |
| InChi Code | InChI=1S/C22H24N2O10/c25-19(26)11-23(12-20(27)28)15-5-1-3-7-17(15)33-9-10-34-18-8-4-2-6-16(18)24(13-21(29)30)14-22(31)32/h1-8H,9-14H2,(H,25,26)(H,27,28)(H,29,30)(H,31,32) |
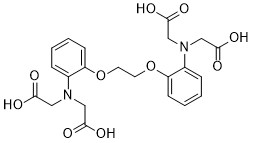
| Chemical Name | 1,2-Bis(2-Aminophenoxy)ethane-N,N,N',N'-tetraacetic acid |
| Synonyms | 85233-19-8; 1,2-Bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid; Bapeta; Ca(II)-bapta; 1,2-Bis(o-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid; UNII-K22DDW77C0; MFCD00036255; |
| HS Tariff Code | 2934.99.9001 |
| Storage |
Powder-20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition | Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs) |
Biological Activity
| Targets | Calcium chelator |
| ln Vitro |
BAPTA (0.3-30 μM; 1 h) inhibits calcium signaling in monodifferentiated NH15-CA2 neuroblastoma and glioma hybrid cells, although it can be employed to stop [Ca2+]-induced cell damage [3]. Phospholipase C (PLC) activity is inhibited by BAPTA (0–10 mM) in a dose-dependent and Ca2+-independent manner [2]. A new family of high-affinity buffers and optical indicators for Ca2+ is rationally designed and synthesized. The parent compound is 1,2-bis(o-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid (BAPTA), a relative of the well-known chelator EGTA [ethylene glycol bis(beta-aminoethyl ether)-N,N,N',N'-tetraacetic acid] in which methylene links between oxygen and nitrogen are replaced by benzene rings. BAPTA and its derivatives share the high (greater than 10(5)) selectivity for Ca2+ over Mg2+ of EGTA but are very much less affected by pH changes and are faster at taking up and releasing Ca2+. The affinity of the parent compound for Ca2+ (dissociation constant 1.1 x 10(-7) M in 0.1 M KCl) may be strengthened or weakened by electron-releasing or -withdrawing substituents on the aromatic rings. The Ca2+ and Mg2+ affinities may further be altered by replacing the ether oxygens by heterocyclic nitrogen atoms. The compounds described are fluorescent Ca2+ indicators absorbing in the ultraviolet region; the very large spectral shifts observed on binding Ca2+ fit the prediction that complexation should hinder the conjugation of the nitrogen lone-pair electrons with the aromatic rings. Derivatives with quinoline nuclei are notable for their high sensitivity of fluorescent quantum yield to the binding of Ca2+ but not of Mg2+. Preliminary biological tests have so far revealed little or no binding to membranes or toxic effects following intracellular microinjection.[1] In cells loaded via patch pipettes with varying concentrations of Ca2+ buffered by 4 mM free BAPTA, light-induced PLC activity, showed an apparent bell-shaped dependence on free Ca2+ (maximum at "100 nM", approximately 10-fold inhibition at <10nM or approximately 1 microM). However, experiments where the total BAPTA concentration was varied whilst free [Ca2+] was maintained constant indicated that inhibition of PLC at higher (>100 nM) nominal Ca2+ concentrations was independent of Ca2+ and due to inhibition by BAPTA itself (IC50 approximately 8 mM). Di-bromo BAPTA (DBB) was yet more potent at inhibiting PLC activity (IC50 approximately 1mM). Both BAPTA and DBB also appeared to induce a modest, but less severe inhibition of basal PLC activity. By contrast, EGTA, failed to inhibit PLC activity when pre-loaded with Ca2+, but like BAPTA, inhibited both basal and light-induced PLC activity when introduced without Ca2+. The results indicate that both BAPTA and DBB inhibit PLC activity independently of their role as Ca2+ chelators, whilst non-physiologically low (<100 nM) levels of Ca2+ suppress both basal and light-induced PLC activity.[2] The effect of the membrane-permeant calcium chelator 1,2-bis-(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid tetra(acetoxymethyl) ester (BAPTA/AM) on ionomycin-induced cellular calcium overload was studied in single differentiated NH15-CA2 neuroblastoma x glioma hybrid cells. To monitor [Ca2+]i we used the fluorescent indicator Fura-2. Preincubation of the cells with 3 microM BAPTA/AM reduced the number of cells showing deregulation of [Ca2+]i during ionomycin-induced calcium influx. The calcium transients elicited by application of KCl were also severely affected by the chelator. These transients, although varying from cell to cell in shape, amplitude and duration, are well reproducible in individual cells. After incubation of cells for 1 h with 0.3-30 microM BAPTA/AM the time course of these cellular transients was markedly slowed. At 1 microM BAPTA/AM, the time constant of decline of [Ca2+]i was increased by a factor of 4.1 +/- 2.4 (n = 14) and the amplitude was reduced to about 50%. With 30 microM BAPTA/AM, the K(+)-induced calcium transients were almost completely inhibited. We conclude that intracellularly loaded calcium chelators may be used for the prevention of [Ca2+]i-induced cell damage, however, at the expense of a disturbed calcium signalling[3]. |
| Cell Assay | Whole-cell recordings. Dissociated ommatidia were prepared from recently eclosed (<2 h) adult flies and transferred to the bottom of a recording chamber on an inverted Nikon Diaphot microscope. As previously described, to avoid series resistance errors, the Kir2.1 current was partially blocked with 4 mM Cs+, which induces a voltage dependent block resulting in a maximum inward current at a voltage of −84 mV. Cells were accurately clamped at this voltage by adjusting the holding potential to find the maximum current, which was regularly checked throughout the recording. The control bath was composed of (in mM): 110 NaCl, 10 KCl, 4 CsCl, 10 N-Tris-(hydroxymethyl)-methyl-2-amino-ethanesulphonic acid (TES), 4 MgCl2, 1.5 CaCl2, 25 proline and 5 alanine. The control intracellular solution was (in mM): 140 K gluconate, 10 TES, 4 Mg ATP, 2 MgCl2, 1 NAD and 0.4 Na GTP. Various buffers (K4BAPTA, K4DBB and K4 EGTA) were made up as 20 or 40 mM stocks, with the same basic intracellular solution but with K gluconate reduced to maintain osmolarity. Two stocks of each buffer were made, one also containing equimolar (20 or 40 mM) CaCl2 and one without Ca2+. These were mixed together with appropriate volumes of the control intracellular solution to generate the various solutions used in this study. The WinMaxC programme was used to calculate the free divalent ion concentration, and pH of all solutions was adjusted to 7.15. To activate reverse Na+/Ca2+ exchange, extracellular NaCl concentration was rapidly reduced by equimolar substitution with LiCl as previously described using a broad-tipped (∼10 μm) puffer pipette, whilst the intracellular solution contained 20 mM Na gluconate and 120 mM K gluconate (otherwise same as control, above). Whole-cell voltage clamp recordings were made using electrodes of resistance ca. 10–15 MΩ, series resistance values were generally below 30 MΩ and were routinely compensated to >80%. Cells were allowed to equilibrate with the patch pipette solution for at least 3 min before making measurements. Data were collected and analyzed at 20 ± 1 °C using Axopatch 1-D or 200 amplifiers and pCLAMP 8 or 9 software. Cells were stimulated by a green light emitting diode (LED), with maximum effective intensity of ca. 4 × 105 photons s−1 per photoreceptor. Relative intensities were calibrated using a linear photodiode and converted to absolute intensities in terms of effectively absorbed photons by counting quantum bumps at low intensities in WT flies, e.g. |
| References |
[1]. New calcium indicators and buffers with high selectivity against magnesium and protons: design, synthesis, and properties of prototype structures. Biochemistry. 1980 May 27;19(11):2396-404. [2]. Inhibition of phospholipase C activity in Drosophila photoreceptors by 1,2-bis(2-aminophenoxy)ethane N,N,N',N'-tetraacetic acid (BAPTA) and di-bromo BAPTA. Cell Calcium. 2005 Dec;38(6):547-56. [3]. Intracellular calcium chelator BAPTA protects cells against toxic calcium overload but also alters physiological calcium responses. Cell Calcium. 1997 Jun;21(6):453-9. |
| Additional Infomation |
BAPTA is a polyamino carboxylic acid in which bis(carboxymethyl)nitrilo groups are bonded to C-2 and C-2' of 1,1'-[ethane-1,2-diylbis(oxy)]dibenzene. It has a role as a chelator. It is a polyamino carboxylic acid and a tetracarboxylic acid. Chelating Agents: Chemicals that bind to and remove ions from solutions. Many chelating agents function through the formation of COORDINATION COMPLEXES with METALS. In conclusion, these results provide compelling evidence that PLC activity in Drosophila photoreceptors is inhibited by BAPTA and DBB. Whilst such non-specific effects of BAPTA are rarely considered, they are not without precedent. For example, it has been reported to inhibit the InsP3 receptor with an IC50 of 340 μM. BAPTA has also been reported to interact with several different calcium-binding proteins including beta-trypsin, parvalbumin, and calmodulin, to inactivate protein kinase C and to act as a blocker of voltage and Ca2+-activated K channels. Where tested, only free BAPTA (as opposed to BAPTA:Ca) appeared to be effective in these instances; the present study is unusual in that the suppression of PLC activity was clearly related to total BAPTA. Although the IC50 (∼8 mM for BAPTA) is rather high, it is not uncommon to use BAPTA at concentrations of 10 mM or more, whilst DBB already has significant potency at 1 mM. Clearly caution needs to be exercised in interpreting results from such experiments, particularly when PLC activity is involved. [2] |
Solubility Data
| Solubility (In Vitro) |
1M NaOH : 33.33 mg/mL (~69.96 mM) DMSO : ~15.62 mg/mL (~32.79 mM) |
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 1.56 mg/mL (3.27 mM) (saturation unknown) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 15.6 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. Solubility in Formulation 2: ≥ 1.56 mg/mL (3.27 mM) (saturation unknown) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 15.6 mg/mL clear DMSO stock solution to 900 μL of 20% SBE-β-CD physiological saline solution and mix evenly. Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. Solubility in Formulation 3: ≥ 1.56 mg/mL (3.27 mM) (saturation unknown) in 10% DMSO + 90% Corn Oil (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 15.6 mg/mL clear DMSO stock solution to 900 μL of corn oil and mix evenly. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.0989 mL | 10.4947 mL | 20.9894 mL | |
| 5 mM | 0.4198 mL | 2.0989 mL | 4.1979 mL | |
| 10 mM | 0.2099 mL | 1.0495 mL | 2.0989 mL |